Representative images of our injection methods for the ZIKV inoculation of embryonic brain are shown in diagrams depicting intracerebral injections (Figure 1A) and intrauterine and intraplacental injections (Figure 1B), illustrating the way the pregnant dam and embryos should be viewed and oriented for surgery (embryonic inoculation protocol). Figure 2A exhibits ZIKV (MEX1-44) infection (immunostained with the antibody against flavivirus group antigen, green) in the E18.5 cerebral cortex. Pax6 (red) labels NPCs in the developing cortex. ZIKV was inoculated into E14.5 embryonic brains. Using the TCID50 assay from tissue samples7, our growth curve analyses show that the ZIKV can efficiently replicate and grow in the developing brains (Figure 2B, 2C), as published7. Figure 2D shows successful infection with DENV2 in the developing cortex using the embryonic inoculation method. DENV2 is detected using an antibody against flavivirus group antigen (green). Figure 2E demonstrates infection with a different strain of ZIKV, the African lineage (MR-766). Figure 2F demonstrates a representative infection for the alternative-route, intraplacental inoculation of ZIKV-Asia (MEX1-44) at stage E10.5.
Figure 3A is a diagram depicting the landmarks used to identify the location of injection into the lateral ventricles for ZIKV inoculation of the P0/1 pups. Figure 3B shows the ZIKV (MEX1-44) infection at P13 cerebral cortex after P0/1 injection. ZIKV (MEX1-44) is detected using antibody against flavivirus group antigen (red). Figure 4 shows the fluorescent beads (red) that were used to practice injection location, and lateral ventricle injection success in adults.
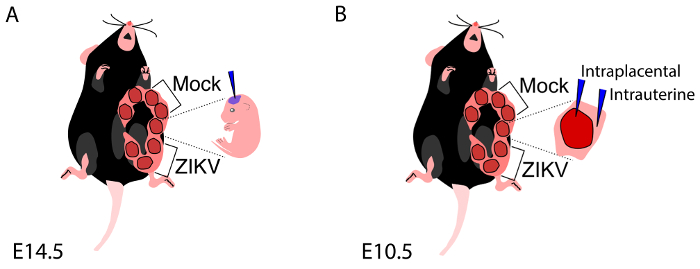
Figure 1: Embryonic inoculation of the ZIKV. (A) Diagram representing exposure of one uterine horn, with a note to avoid injection of embryos adjacent to vagina and the most lateral embryo along the horn. (B) Diagram representing exposure of one uterine horn, depicting the location of intraplacental and intrauterine injections. These diagrams have been modified from their original publication21. Please click here to view a larger version of this figure.
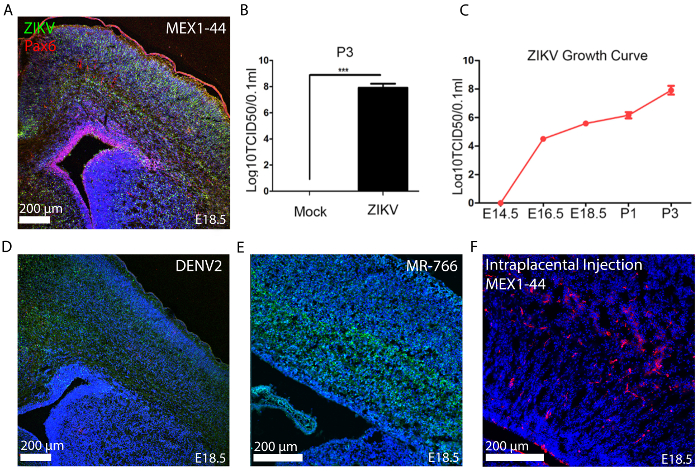
Figure 2: ZIKV Embryonic Inoculation at E14.5. (A) At E18.5 the embryonic brain is infected with ZIKV-Asia (MEX1-44) across all layers of the developing neocortex (scale bar is 200 µm). Neural progenitor cells are immunolabeled with Pax6 (red) and ZIKV via flavivirus antigen (green). (B) TCID50 results from Postnatal Day 3 (P3) brain tissue inoculated at E14.5 with ZIKV-Asia (MEX1-44). Error bars indicate s.e.m. of three independent measurements with one mock and one ZIKV-infected brain in each measurement (***p <0.0001, Student's t-test)7. (C) TCID50 results describing typical increased growth curve of ZIKV viremia in infected fetal brain tissue. Error bars indicate the s.e.m. of three independent measurements with one mock and one ZIKV-Asia (MEX1-44) infected brain in each measurement. Analysis of variance (ANOVA) detects a significant increase in viral titer as development proceeds7. (D) Dengue virus (DENV2) infected representative histology (E14.5 embryos inoculated with ~ 1 µL of 3.4 x 105 TCID50/mL, scale bar is 200 um) and (E) ZIKV-Africa (MR766) infected representative histology (scale bar 200 µm). (F) ZIKV-Asia (MEX1-44) infected representative histology from the intraplacental injection strategy (scale bar is 200 µm). In all images, Hoechst stains nuclei. These figures have been modified from their original publication7. Please click here to view a larger version of this figure.
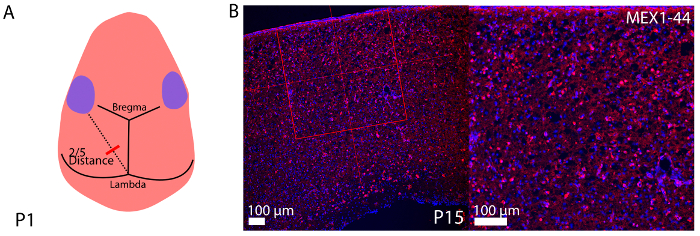
Figure 3: Representative histology of P1 inoculation of ZIKV. (A) Diagram demonstrating the method to determine the injection location into the lateral ventricles of P1 pups. (B) Representative coronal cryosections immunostained for flavivirus group antigen with low (on left, scale bar is 100 µm) and high (on right, scale bar is 50 µm) magnification of P1 inoculation of ZIKV at P15 (~ 1 μl 3.4 x 105 TCID50/mL ZIKV). Hoechst stains nuclei. Please click here to view a larger version of this figure.
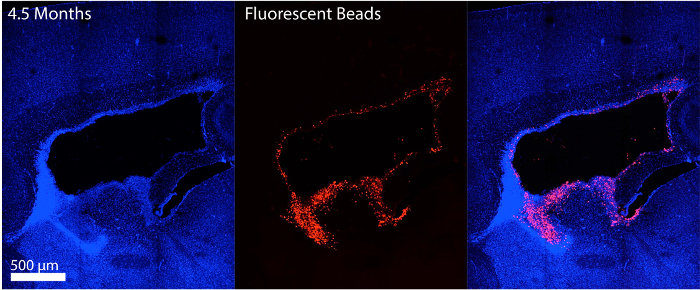
Figure 4: Adult stage intraventricular injection. Adult mouse injected with fluorescent beads was sacrificed shortly after surgery to determine the injection location success (scale bar is 200 µm). Sagittal cryosection could be viewed immediately after sectioning as beads do not require further staining. Hoechst stains nuclei. Please click here to view a larger version of this figure.
