A pathogenesis assay (Figure 1) using C. albicans and C. elegans has previously been described by our lab17,18 and other labs19,20. We demonstrate the amenability of using C. elegans to study C. albicans virulence showing that C. albicans cells are quickly ingested by the worms and accumulate in the intestinal lumen causing slower locomotion, deformed anal region (Dar) (Figure 2A), swelling of the vulva (Figure 2B), distention of the intestine (Figure 3A), and lethality (Figure 3, Figure 4). We also measure the life span of infected worms as a quantitative measure of fitness. For example, C. elegans infected with C. albicans live 10-12 days compared to 20-22 for uninfected controls. In C. elegans infected with the C. albicans double knock out mutant, efg1/efg1 cph1/cph1, which are two key virulence factors required for infection in mouse, rats and human epithelial models21,23, nematodes survived significantly longer than controls (Figure 3B). These experiments suggest that some of the lessons we learn in this simpler model about C. albicans virulence may remain valid in higher mammals and vice versa.
We also show that the nematode model system can be pharmacologically modulated (Figure 3B). In the presence of fluconazole, the most commonly prescribed antifungal drug, worms challenged with C. albicans survived significantly longer than controls. This proof of principle experiment suggests that the nematode model can be used for small molecule screening. Indeed, our C. elegans model was instrumental in identifying filastatin, a small molecule inhibiting various aspects of fungal virulence24.
Disease phenotypes and microscopic analysis of C. elegans infection
C. elegans were exposed to C. albicans over a period of six days and observed for signs of infection, progression of disease, and death. The Dar phenotype is most visible by day 4 of the survival assay, as noted by a protruding anal region that is not visible in the uninfected animal (Figure 2A). Worms infected by C. albicans are also known to exhibit swelling in the vulva region (Figure 2B). In both cases, the worm is unable to clear the infection after reaching this stage. In order to visualize colonization of C. albicans in the intestinal lumen, worms were fed wild-type C. albicans tagged with RFP, which cause areas of colonization to fluoresce red (Figure 3A). In order to produce these images, worms were anesthetized on a 2% agarose pad containing 0.01 M sodium azide. Worms were exposed to either wild-type C. albicans or RFP-tagged C. albicans, and transferred into 5 µM M9 buffer on the agarose pad. The agarose pad was then covered with a coverslip. Worms were viewed at 200X and 400X magnification using an inverted microscope with fluorescent microscopy capabilities. Images were created under differential interference contrast (Nomarski) and epifluorescence optics. The fluorescent images were enhanced using software (Figure 3A). Time series micrographs of infected worms determined that C. albicans colonized the intestinal lumen by the third day of the assay17. Infected worms showed more severe intestinal distension than observed in the uninfected control.
Genetic and pharmacological tools to study infection
Next, we tested the model using genetic and pharmacological modulation. We also tested previously documented virulence factors that regulate hyphal transition of C. albicans25 and have been shown to be important in in vivo infections of mice and nematodes20,26. We tested the ability of the worms to survive infection caused by C. albicans efg1/efg1 cph1/cph1 double mutant. Efg1 is a highly-conserved transcription factor and an essential component of the cyclic AMP/protein kinase A (PKA) metabolic pathway. In C. albicans this pathway regulates hyphal morphogenesis27, white-opaque switching28, and an arsenal of key virulence factors. These virulence factors include Hwp1 and Hwp2, two yeast-specific cell wall proteins involved in adhesion and biofilm formation, Eap1, a cell-wall adhesin involved in binding to human epithelial cells29, and Sap5, a hydrolytic enzyme involved in epithelial tissue invasion30. Cph1 is a transcription factor that regulates many metabolic processes including mating, filamentation, and biofilm formation and has been shown to play a critical role in damaging epithelial cells31 and human reconstructed epithelium21. Disruption of either of these genes has a significant impact on virulence and simultaneous disruption of both in cph1/cph1efg1/efg1 results in dramatic virulence reduction in various animal models including mice25, Drosophila32, zebrafish33,34 and moth34. The cph1/cph1efg1/efg1 double mutant is considered by the C. albicans community as the avirulent strain by definition and the gold standard for validation of novel model systems. The efg1Δ and cph1Δ single mutants showed decreased Dar (~10% and ~50%, respectively) compared with the cognate wild type, while the efg1Δ cph1Δ double mutant failed to elicit the Dar response17. These results recapitulate the pattern of virulence in mice, where the cph1∆ mutant is slightly attenuated, the efg1∆ mutant is significantly attenuated, and the double mutant is completely avirulent25. Worms infected with cph1/cph1 efg1/efg1C. albicans double mutant lived statistically significantly longer than controls (p <0.01 for both Log Rank and Breslow statistical test, n=60) suggesting that these two genes are required for C. albicans virulence against C. elegans (Figure 3B).
In order to explore the possibility of using our C. elegans model for potential drug screening, we tested the effect of the addition of fluconazole on the ultimate outcome of the infection. We used a variety of different concentrations of fluconazole and found that 50 µM gave us the most significant results. Worms infected with C. albicans and 50 µM fluconazole (Figure 3B) lived statistically significantly longer than controls (p<0.01 at both Log Rank and Breslow statistical test, n=60). This concentration was empirically determined (see the note at the end of protocol step 7, "infection plate set up"). These proof of principle experiments showed that our nematode model can be used for small molecule antifungal screening. The model was in fact instrumental in the discovery of filastatin, a small molecule inhibiting various aspects of fungal virulence that is currently undergoing further preclinical studies24. Accordingly, our assay is suitable for exploring the virulence strategies of C. albicans and pharmacological agents.
Study of innate host response
Next, we wanted to study the reciprocal host defenses against pathogens, since aspects of this innate immunity are conserved in mammals11,35. It is well known that C. elegans produces ROS upon both bacterial and fungal infections18,36,37 as part of its defense mechanism. ROS have a biocidal effect on invading organisms and play a major role in innate immunity. zcf15/zcf15 hyper susceptibility to ROS in vitro led us to hypothesize that its reduced virulence in C. elegans was due to a reduced ability to withstand the host's generated ROS. To test our hypothesis, we determined the ability of zcf15/zcf15 to kill either wild-type worms or worms with an impaired ability to produce ROS. Representative C. albicans mutant zcf15 that is sensitive to reactive oxygen species (Figure 4A) showed reduced virulence in wild-type C. elegans. C. elegans responds to ingestion of pathogen by producing extracellular ROS in the intestinal lumen via Ce-Duox1, an NADPH oxidase coded by the gene bli-3. During ROS production, the intestinal cells also produce intracellular antioxidants via DAF-16 to protect its own tissues from the ROS damaging effects36,38. Ce-Duox1 is a protein with an N-terminal peroxidase domain, a C-terminal superoxide-generating NADPH-oxidase domain and two central calmodulin-binding sites39. Upon microbial infection, this protein uses cytosolic NADPH to generate extracellular toxic ROS in the intestinal lumen to counteract the infection. Throughout a series of biochemical assays in a 2009 study, Jain et al. showed that ROS are abundantly produced upon yeast infection and that bli-3 loss of function via bli-3(e767) dramatically reduces the ability of nematodes to produce ROS. As shown in Figure 4B, worms challenged with zcf15/zcf15 survived significantly longer than wild type or complemented strains (p <0.01 by the log-rank test) indicating that ZCF15 is required for virulence. However, when we challenged ROS-deficient C. elegansbli-3(e767), we obtained kinetics of killings that were comparable between zcf15/zcf15, wild type and complemented strain (Figure 4C). This evidence indicates that zcf15/zcf15 fail to kill nematodes unless the host's ability to produce ROS is compromised. Taken together, these results suggest that ZCF15 is required for full virulence and that this gene is likely involved in the pathogen's ability to resist host generated ROS. To the best of our knowledge, this is the first time that ZCF15 has been shown to be involved in pathogenicity and ROS resistance.
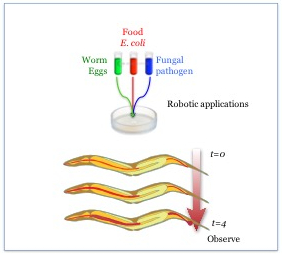
Figure 1: A schematic representation of C. elegans infection. C. elegans are exposed to a mix of E. coli strain OP50 and a pathogen and observed over a period of 4 days after reaching adulthood for signs of infection and progression of disease.
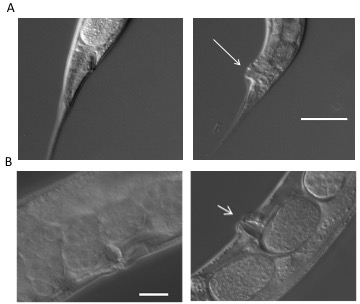
Figure 2: Disease phenotypes. (A) Deformed anal region (Dar) is visible (indicated with an arrow) as a swelling in the post anal region of infected worms (right panel) four days post exposure. The worms on the left panel are uninfected and serve as a control. (B) A subset (~15%) of infected worms show a swelling of the vulva (right panel, indicated with an arrow) compared to uninfected control worms (left panel) that represents disseminated infection resulting in matricidal death. All images were taken as described in the representative results18.
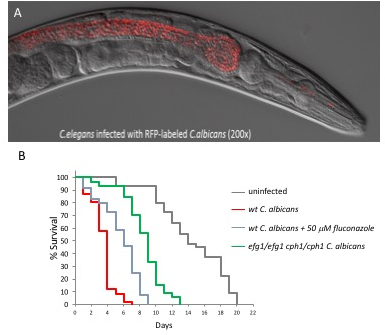
Figure 3: The C. albicans – C. elegans model is amenable to microscopic manipulation, and can be pharmacologically and genetically modulated. (A) C. albicans tagged with RFP accumulation in the nematode intestinal lumen day 3 post infection. Yeast cells are quickly ingested by the worms and accumulate in the intestinal lumen completely intact, indicating that they are able to survive the mechanical crushing of the pharynx. Images were taken as described in the representative results18. (B) Kaplan-Meier survival curves of nematodes challenged with wild-type C. albicans, cph1/cph1efg1/efg1 double mutant or wild-type C. albicans + 50 µM of fluconazole compared to uninfected control worms.
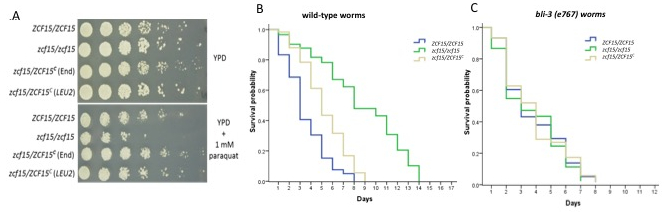
Figure 4. ZCF15 is required to withstanding C. elegans generated ROS. (A) ZCF15 is responsible for wild type resistance to paraquat, which is known to generate reactive oxygen species. Wild type, knockout or complemented strains were grown in liquid culture overnight and resuspended to OD=1. Cultures were each diluted 1:5 and plated onto YPD or YPD containing 1 mM paraquat. C. elegans produce ROS via bli-3 in order to combat pathogens that invade the intestinal lumen38. (B) Kaplan-Meier survival curves show that ZCF15 deletion limits the killing of wild-type worms. (C) The Kaplan-Meier survival curve shows similar rates of survival between zcf15/zcf15, wild type, and complemented strain when ROS-deficient C. elegansbli-3(e767) were infected.
