Figure 1 illustrates a schematic time table for metabolic phenotyping of mice on diets. At an age of approximately 6 weeks, mice should be placed on an HFD, while an LFD-group may serve as the control group. Importantly, body weight should be determined weekly to observe if there is an expected increase in body weight. Any kind of stress (e.g., noise or aggressive male behavior) can interfere with body weight gain and should be eliminated immediately. Each cohort of mice for diet experiments should consist of at least 10 mice because these diet-experiments are time-consuming, and outliers are frequent (e.g., mice not gaining weight or mice with abnormal glucose or insulin levels). After the selected period of time (depending on the study hypothesis and the time point of expected changes), OGTT and ITT can be performed for the evaluation of glucose tolerance and insulin action. In this paper, late time points for the metabolic test were chosen.
Importantly, there should be a recovery time of at least 1 week between the OGTT and ITT as these experiments lead to substantial blood-loss and are thus very stressful for mice. If the blood collection volumes are decreased (e.g., if performing an ITT without additional blood collection first), this recovery period may also be shortened or omitted, in line with the guidelines for multiple blood draws in animals14,15,16,17.
In this large study with 60 C57BL/6J mice in total, half of the mice were set on HFD or LFD at an age of 6 weeks (n = 30/group) and body weight gain was monitored for 16 weeks on diet. The consumption of HFD resulted in a significant increase in body weight as shown in Figure 4. At 6 weeks of age, the body weight was 20.2 g in both groups. Whereas mice on LFD showed a consistent, slightly increasing body weight (31.2 g ± 2.7) during the observed period, mice on HFD increased their body weight rapidly, especially during the first weeks and reached their body weight maximum after 16 weeks on the diet. Although the weight curves showed a similar pattern during the experiment, the mice of the HFD-group reached a 1.5- to 2-fold higher body weight (44.4 g ± 4.0) compared to the LFD-fed mice.
To investigate the metabolic phenotype of the two cohorts, an OGTT (Figure 5) and ITT (Figure 6) were performed. As blood volume is limited in small rodents, a point-of-care (POC) assay for diabetic humans (glucometer) was used to monitor blood glucose levels during these metabolic phenotyping experiments. As demonstrated in Figure 2, the blood glucose monitors are easy to use, need only a small drop of blood, and display blood glucose levels within seconds for documentation. Figure 5 presents the time course of absolute glucose (Figure 5a–b) and absolute insulin (Figure 5c) levels during the OGTT. Generally, a healthy mouse with normal glucose tolerance shows a characteristic rapid rise in blood glucose, reaching its peak 15-30 min after the glucose challenge.
Subsequent glucose uptake, primarily conducted by muscle, fat-tissue, and liver-tissue leads to a gradual decrease of the blood glucose concentration. In all experiments, the LFD-fed mice served as the glucose tolerant control group and therefore fulfilled the expected metabolic profile: the peak of the blood glucose levels of ~240 mg/dL was reached approximately 15 min after glucose administration, immediately followed by a decrease reaching basal levels approximately 60 min after the glucose challenge, indicating proper glucose elimination. In sharp contrast, HFD-mice peaked at approximately ~320 mg/dL glucose and showed nearly no disposal of glucose, indicating glucose resistance. When blood glucose levels between two groups already differ in the fasting state (as in this representative example), a calculation of the area under the curve (AUC) above baseline glucose should be performed to validate the results (Figure 5a–b).
Additionally, the circulating blood insulin levels were determined using an insulin-ELISA test (Figure 5c) in order to provide more information about the underlying pathophysiology in this model. Whereas the insulin levels were nearly unchanged in the control group, mice fed an HFD showed 16-fold elevated fasting levels compared to the control group, as well as a greatly increased insulin response, indicating HFD-induced compensatory hyperinsulinemia as an attempt to counterbalance decreased glucose elimination capacity, which may be caused by insulin resistance. However, be aware not to over-interpret the results of the OGTT, as this test does not directly evaluate insulin action and should not be used to conclude statements about insulin resistance.
To measure insulin sensitivity in the HFD-fed mice, an ITT was performed 1 week after OGTT (Figure 6a). In this assay, the degree to which blood glucose concentrations fall following insulin administration represent the efficiency of whole-body insulin action. The HFD-fed mice showed an impaired reduction of blood glucose levels compared to the LFD-fed control group, at all time points during the ITT, thus suggesting insulin resistance. The ITT results are usually presented as the time course of glucose levels, but additionally also the inverse AUC below baseline glucose may be shown as demonstrated in Figure 6b. If the groups which are compared have similar fasting blood glucose levels (which is not the case in this experiment), the glucose levels during ITT can also be presented as the percentage of basal glucose. As in mice, a counter-regulatory response to insulin is activated if the blood glucose levels fall below ~80 mg/dL18: defects in this counter-regulatory response in a particular mouse model may be misinterpreted as an increase in insulin sensitivity. During HFDs and subsequent metabolic phenotypic experiments, outliers may frequently occur. Mice which do not gain weight on HFD, or those showing abnormal fasting glucose and/or insulin levels should be excluded from the analysis. For the latter two, an outlier test may be performed for each experimental group separately (e.g., Grubbs test)
In this study, as an example we showed and interpreted data of metabolic experiments in vivo, carried out on mice with diet-induced obesity, glucose intolerance, and insulin resistance, and compared them to a control group with normal body weight. As expected, there was impaired glucose tolerance and hyperinsulinemia in obese mice consistent with insulin resistance compared to the age-matched control mice; this was uncovered using well-established, reliable, time- and budget-friendly methods, which are relatively easy to perform. Differences in glucose tolerance, insulin levels as well as in insulin sensitivity, which are all obtained by the presented methods of OGTT and ITT, can often help to plan the next steps of a study, which may include more sophisticated experiments such as hyperglycemic or hyperinsulinemic clamps, as well as experiments with isolated pancreatic islets.
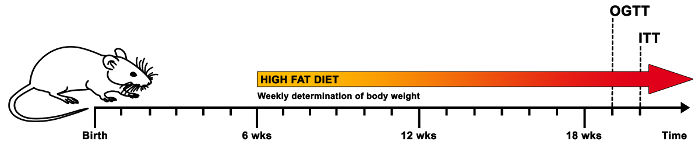
Figure 1. Schematic time table for a suggested diet regime and metabolic experiments in vivo. In order to investigate the metabolic effects of HFD in mice, the animals of the experimental group are placed on HFD at approximately 6 weeks of age, while the control group receives an LFD. The body weight of the mice should be determined on a weekly basis to assess proper weight gain. After approximately 12 weeks on diet (or a selected time point depending on the research hypothesis), the metabolic phenotype of the mice is evaluated by an OGTT followed by 1 week of recovery-time and subsequently an ITT. Please click here to view a larger version of this figure.

Figure 2. Methods for blood sampling during metabolic experiments. For the OGTT as well as for the ITT, where repeated blood sampling is required, we recommend drawing blood via carefully cutting a 1-2 mm piece of the tail tip with sharp scissors (Variant A), followed by the determination of blood glucose levels with a glucometer and further collection of blood with a capillary to determine insulin levels and other relevant blood values. Alternatively, blood might also be sampled via the tail vein (Variant B) or by arterial catheterization (not shown). Please click here to view a larger version of this figure.
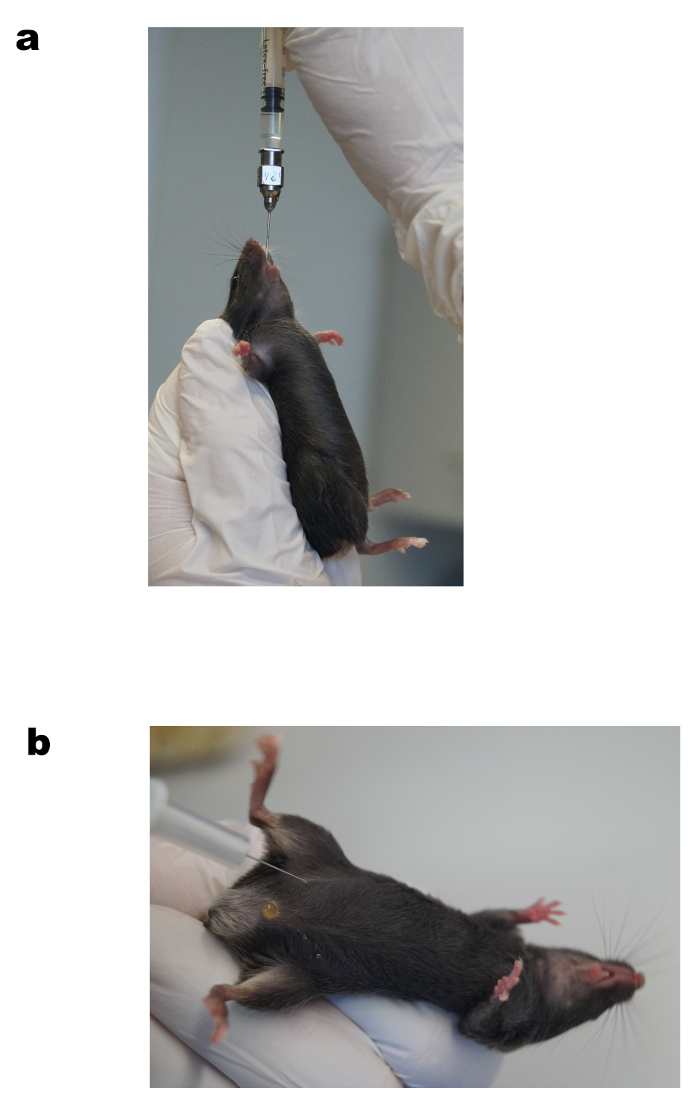
Figure 3. Oral gavage of glucose (a) and intraperitoneal insulin injection (b). Representative images of oral glucose administration using a feeding needle during OGTT (a) and the intraperitoneal injection of insulin during ITT (b). See protocol for a detailed description. Please click here to view a larger version of this figure.
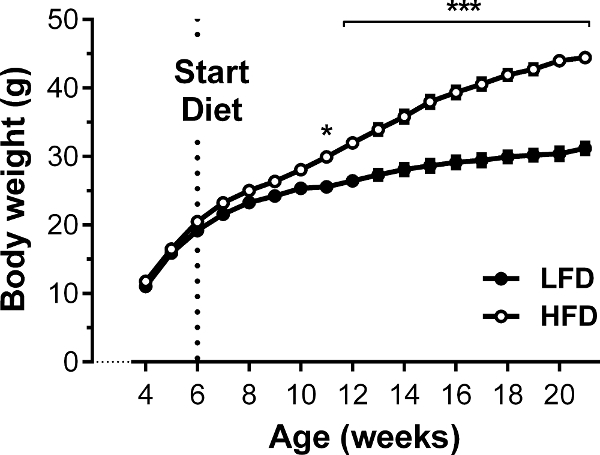
Figure 4. Body weight gain of HFD-fed and LFD-fed C57BL/6J mice. C57BL/6J mice were either set on 60% HFD, or 10% LFD to serve as a control, for a period of 20 weeks. Whereas mice on HFD showed an expected increase in body weight, especially in the first weeks on diet, LFD-fed mice showed nearly constant body weight during the observed period. Results are mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001. n = 30 per group. ANOVA and Tukey's post hoc test were used to test for differences. Please click here to view a larger version of this figure.
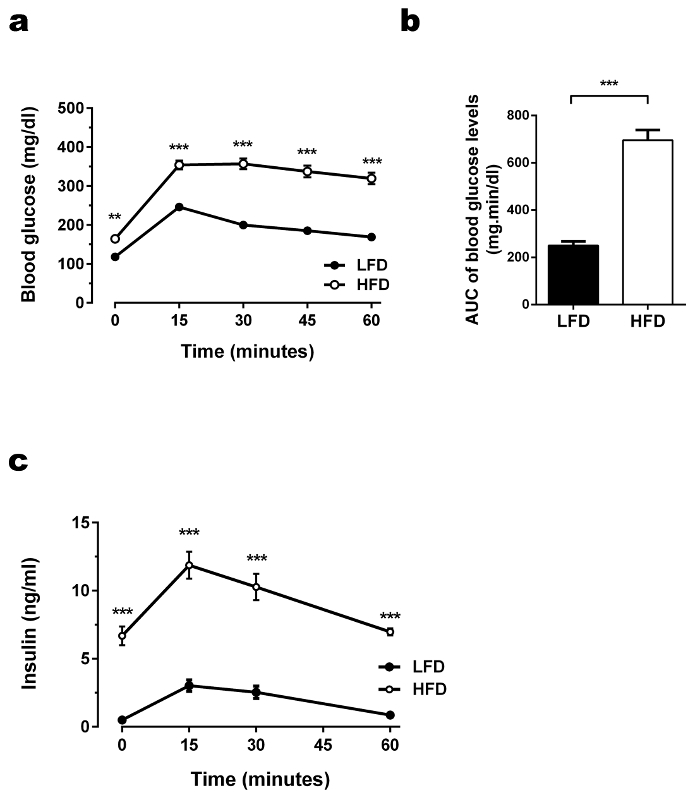
Figure 5. OGTT performed in HFD-fed and LFD-fed C57BL/6J animals. (a) Glucose levels during OGTT. After an overnight fast, glucose (mg/dL) levels were measured in fasting state and 15, 30, 45, and 60 min after administering glucose solution orally via gavage (1 g glucose/kg). Glucose levels in the HFD-group were elevated in the fasting state as well as after glucose challenge. The increase reached its peak after 15 min followed by a delayed and slow decrease. Results are mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001. n = 30 per group. Statistical analysis was performed using ANOVA and Tukey's post hoc test.(b) Glucose area under the curve (AUC) during OGTT. To calculate the baseline corrected AUC, basal glucose levels (time point 0) were subtracted from all later obtained blood glucose levels for each mouse individually, followed by the calculation of the individual AUCs. The AUC above the baseline glucose illustrates the glucose resistance in the HFD-fed mice. Statistical analysis was performed using ANOVA and Tukey's post hoc test (glucose levels) or Student's two tailed t-test (AUC). (c)Insulin levels during OGTT. Insulin (ng/mL) levels were measured after a fasting period of 4 h and 15, 30, and 60 min after administering glucose solution orally via gavage (1 g glucose/kg). HFD-fed mice not only compensated for the glucose injection with a higher increase in blood insulin levels, they also started and ended the OGTT with elevated insulin levels compared to the control group. Results are mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001. n = 30 per group. Statistical analysis was performed using ANOVA and Tukey's post hoc test. Please click here to view a larger version of this figure.
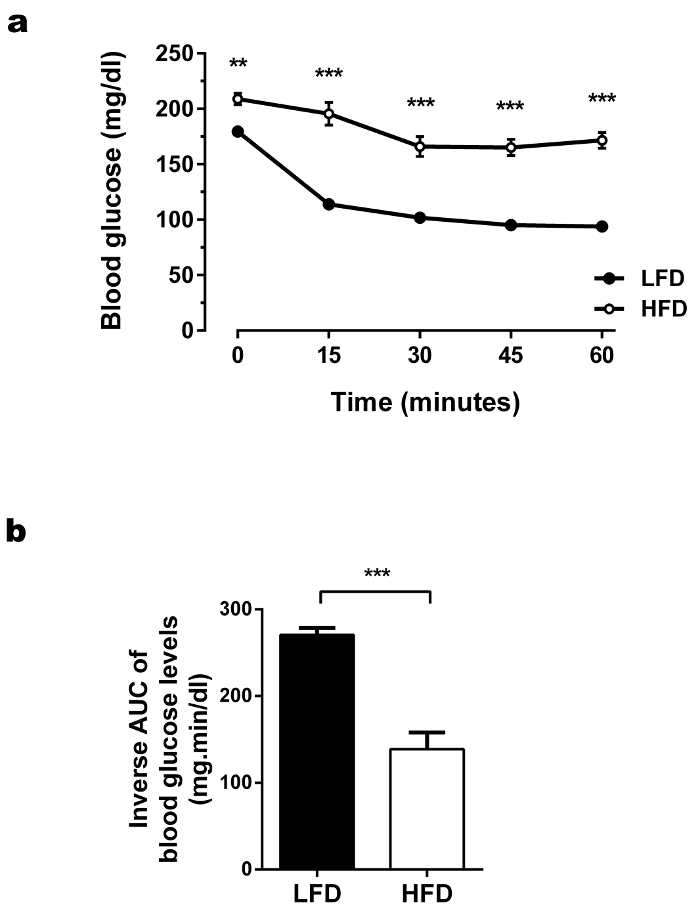
Figure 6. ITTs performed in HFD-fed and LFD-fed C57BL/6J animals. (a) Glucose levels during ITT. The glucose (mg/dL) levels were measured in fasting state and 15, 30, 45, and 60 min after injection of insulin intraperitoneally (0.75 U insulin/kg). During ITT, HFD-fed mice showed elevated glucose levels. The blood glucose levels were not adequately lowered in the HFD-fed mice after insulin injection. Results are mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001. n = 30 per group. Statistical analysis was performed using ANOVA and Tukey's post hoc test. (b) Glucose area under the curve (AUC) during ITT. To calculate the baseline corrected inverse AUC, basal glucose levels (time point 0) were subtracted from all later obtained blood glucose levels for each mouse individually. The values were inverted (multiplication with -1), followed by the calculation of the individual AUCs. As a consequence of the higher glucose level in HFD-fed mice during OGTT, the baseline corrected inverse AUC was lower in the HFD-fed mice compared to control mice, which further suggested decreased insulin sensitivity. Statistical analysis was performed using ANOVA and Tukey's post hoc test (glucose levels) or Student's two tailed t-test (inverse AUC). Please click here to view a larger version of this figure.
Supplementary Figure 1. Checklist for experiment preparation. Please click here to download this file.
Supplementary Figure 2. Insulin levels during ITTs. Plasma insulin levels during the ITT in the LFD-fed versus the HFD-fed groups showed similar dynamics in the plasma insulin levels after insulin injection in both groups. As expected, the HFD mice exhibited strongly increased basal insulin levels compared to the control group. Further, the increase in the insulin levels in the HFD-fed mice was stronger, which may be partially caused by the overestimation of lean body mass if the amount of injected insulin is calculated based on the whole-body mass (the conventional normalization approach) as performed in this experiment. However, the insulin response was impaired in the HFD-fed group (insufficient reduction of plasma glucose levels), thus further emphasizing the insulin resistant state in these animals. Results are mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001. Statistical analysis was performed using ANOVA and Tukey's post hoc test. Please click here to download this file.
