Mange proteiner opfylde deres funktion i dynamisk komplekser med andre molekyler, medieret af konformationelle ændringer og forbigående foreninger på en bred vifte af tidsskalaer1,2,3. Koblet til en ekstern energikilde (fx ATP) disse dynamiske interaktioner kan føre til tekstretning i en funktionel cyklus og i sidste ende bevare ikke-ligevægt steady-state i en celle, en forudsætning for liv.
For fuldt ud at forstå disse molekylære maskiner, er en statisk beskrivelse styret af strukturelle undersøgelser ikke tilstrækkelig. Derudover er det vigtigt at have kendskab til de underliggende kinetiske model og at bestemme de kinetiske sats konstanter. Flere eksisterende metoder giver forskere til at studere dynamikken i binære interaktioner mellem to molekyler af interesse, fx overflade plasmon resonans, afslapning metoder med en spektroskopiske udlæsning (fx hop eller stoppet-flow teknikker), og Kernemagnetisk resonans. Deres anvendelighed er imidlertid i de fleste tilfælde begrænset til enkle to-stats systemer (f.eks. en bundet og en ubundet stat) på grund af den gennemsnit iboende til bulk eksperimenter. I tilfælde hvor flere stater eller mellemprodukter er involveret, giver de kun en kompleks blanding af sats-konstanter. Enkelt-molekyle metoder såsom optiske eller magnetisk pincet eller to-farve smFRET, dvs., én donor og en acceptor fluorophore, med en overflade-immobiliseret prøve kan gendanne sats konstanter for alle observerede konformationelle ændringer. Men når det kommer til interaktioner, der berører mere end én bindingssted, disse metoder være begrænset og oplysninger om mulige korrelationen mellem to (eller flere) interaktioner vil kun være tilgængelig via indirekte konklusioner fra et sæt af forsøg.
Multi-farve smFRET4,5,6,7,8,9 tilbyder mulighed for at studere samspillet mellem disse komponenter direkte, i realtid og under i nærheden af fysiologiske forhold10. Dette gør det muligt at undersøge eksempelvis strukturafhængigt bindingen af en ligand eller et andet protein8,9,11. Den samlede tilgang præsenteres her er at mærke de(n) af interesse på bestemte holdninger, til at vedhæfte en protein til overfladen af måling kammer og spore fluorescens-intensiteten over tid på en prisme-type TIRFM (for detaljer se 9 , 12). den geografiske nærhed af de forskellige farvestoffer kan derefter bestemmes fra energioverførsel mellem dem. Mærkning strategier kan variere fra protein til protein (gennemgik i 13) og retningslinjer for at undgå artefakter i smFRET målinger findes14.
Da en donor farvestof kan overføre energi til forskellige acceptor farvestoffer i en multi-farve smFRET eksperiment, er den relative placering af alle farvestoffer ikke tilgængeligt fra excitation af et farvestof alene15,16. Men i kombination med skiftevis laser excitation (ALEX17, og gennemgik i 18) denne metode giver alle spatio-temporale oplysninger på lynhurtige og sub nanometer opløsning.
Hovedstol, høj opløsning strukturelle oplysninger kan opnås ved hjælp af Inter farvestof afstande beregnes ud fra kombinationen af alle fluorescens intensiteter i en multi-farve smFRET eksperiment med ALEX. Men her vi fokusere på staten identifikation og adskillelse samt udvinding af kinetiske modeller, hvor multi-farve smFRET er uundværlig. Når “kun” struktur bestemmelse af triangulering ønskes, kan en række enklere tofarvet smFRET eksperimenter med høj signal-støj-forholdet være udført12,19.
Vi bruger den delvise fluorescens (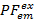 ) som en proxy for energioverførsel mellem to fluorophores7. PF er beregnet ud fra fluorescens intensitet svarer til FRET effektiviteten af en to-farvet eksperiment:
) som en proxy for energioverførsel mellem to fluorophores7. PF er beregnet ud fra fluorescens intensitet svarer til FRET effektiviteten af en to-farvet eksperiment:
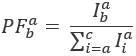
Hvor 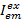 er intensiteten i emission kanal em efter excitation med farve ex, og c er acceptor med den længste bølgelængde. Opdagelse kanaler repræsenterer den samme holdning i prøve salen men optage forskellige spektrale intervaller lysets fluorescens. Den samme id for excitations- og anvendes i denne protokol (dvs., “blå”, “grønne” og “røde”).
er intensiteten i emission kanal em efter excitation med farve ex, og c er acceptor med den længste bølgelængde. Opdagelse kanaler repræsenterer den samme holdning i prøve salen men optage forskellige spektrale intervaller lysets fluorescens. Den samme id for excitations- og anvendes i denne protokol (dvs., “blå”, “grønne” og “røde”).
På grund af eksperimentelle mangler afhænger de målte fluorescens intensiteter ikke kun om energioverførsel, men også på fluorophore og setup egenskaber. For at opnå den sande energieffektivitet overførsel mellem to fluorophores, skal de målte intensitet korrigeres. Den følgende procedure er baseret på reference9. Korrektionsfaktorer for tilsyneladende lækage (lk, dvs, påvisning af fotoner fra en fluorophore i en kanal udpeget til en anden farve) og tilsyneladende gamma (ag, dvs., fluorescens kvanteudbytte af farvestoffet og den opdagelse effektivitet af kanalen) er fremstillet af enkelt-molekyle spor, der viser en acceptor blegning begivenhed.
Udsivning af donor farvestof i alle mulige acceptor kanal er beregnet ud fra alle datapunkter i de registrerede fluorescens spor hvor acceptor farvestof bleget men donor er stadig fluorescerende (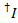 ):
):
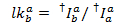
Medianværdien af lækage histogrammet bruges som tilsyneladende lækage faktor. Efter korrektion for lækage bestemmes den tilsyneladende gamma faktor fra det samme sæt af spor. Det beregnes ved at dividere ændringen af fluorescens i acceptoren kanal af ændringen af fluorescens i donor kanal ved blegning af acceptoren farvestof:
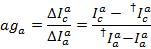
Hvor c er igen den opdagelse kanal for acceptor med den længste bølgelængde. Medianen af den resulterende distribution anvendes som tilsyneladende korrektionsfaktoren.
De korrigerede intensiteter i hver kanal er fremstillet af:
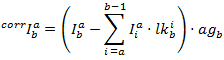
PF er derefter beregnes i henhold til:
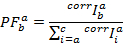
Forskellige populationer kan adskilles i multi-dimensionelle rum som PFs. Position og bredde i hver stat bestemmes af montering af data med multi-dimensional Gaussisk funktioner. Efterfølgende optimering af en global HMM baseret på alle PF spor giver en kvantitativ beskrivelse af de observerede kinetik. Selv små ændringer i satserne kan registreres.
HMMs give en måde at udlede en stat model fra en samling af støjende tid spor. Systemet anses for at være i en af et sæt af diskret, skjult stater på et givet tidspunkt og den faktiske observation (dvs. emissionen) er en probabilistisk funktion af denne skjulte tilstand20. I tilfælde af TIRFM smFRET data, kan den emission Sandsynligheder bjeg pr. stat, jeg modelleret af kontinuerlig Gaussisk betafordelingen funktioner. På regelmæssigt fordelte diskret tid point, kan overgange fra den ene til en anden stat forekomme efter overgangen sandsynligheden af, er tid-invariant og kun afhænger af den aktuelle tilstand. Overgangen matrix A indeholder disse overgang sandsynligheder enij mellem alle skjulte stater. Starttilstand fordeling 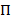 giver de statsspecifikke sandsynligheder
giver de statsspecifikke sandsynligheder 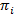 for den første tidspunkt for en tid spor. Bruger en maksimum-sandsynligheden tilgang, kan disse parametre være optimeret til at bedst beskriver dataene med den frem-tilbage og Baum-Welch algoritmer20,21. Dette giver maksimal sandsynligheden estimatorer (MLE). Endelig, den stat sekvens, der mest sandsynligt produceret bane af observationer kan udledes med Viterbi algoritme. I modsætning til andre HMM analyser af smFRET data24,25,26 bruger vi ikke HMM som en blot “udjævning” af data, men ekstrakt kinetic state-modellen fra datasættet uden behov for passende hviletid histogrammer27. HMM analyse er færdig med in-house scripts ved hjælp af Igor Pro. Gennemførelsen af kodeksen er baseret på reference21. Vi leverer en software kit og eksemplarisk data på vores webside for at følge afsnit 5 og 6 i denne protokol (https://www.singlemolecule.uni-freiburg.de/software/3d-fret). Fuld software er tilgængelig efter anmodning.
for den første tidspunkt for en tid spor. Bruger en maksimum-sandsynligheden tilgang, kan disse parametre være optimeret til at bedst beskriver dataene med den frem-tilbage og Baum-Welch algoritmer20,21. Dette giver maksimal sandsynligheden estimatorer (MLE). Endelig, den stat sekvens, der mest sandsynligt produceret bane af observationer kan udledes med Viterbi algoritme. I modsætning til andre HMM analyser af smFRET data24,25,26 bruger vi ikke HMM som en blot “udjævning” af data, men ekstrakt kinetic state-modellen fra datasættet uden behov for passende hviletid histogrammer27. HMM analyse er færdig med in-house scripts ved hjælp af Igor Pro. Gennemførelsen af kodeksen er baseret på reference21. Vi leverer en software kit og eksemplarisk data på vores webside for at følge afsnit 5 og 6 i denne protokol (https://www.singlemolecule.uni-freiburg.de/software/3d-fret). Fuld software er tilgængelig efter anmodning.
Tid punkter i data med PF < -1 eller PF > 2 i enhver opdagelse kanal er tildelt minimal emission sandsynligheden for alle stater (10-200). Dette forhindrer kunstige overgange på disse datapunkter.
Parametre for emission Sandsynligheder er fremstillet af anfald af 3D PF histogrammet med Gaussisk funktion som beskrevet i trin 5.7. Disse parametre holdes fast under optimering af HMM.
I metoden præsenteres starttilstand distribution vektor og overgangen matrix anvendes globalt til at beskrive hele ensemblet af spor. De opdateres baseret på alle N molekyler fra datasæt efter reference27.
Startparametre i den oprindelige tilstand distribution bestemmes ud fra 2D fremskrivninger af PF histogram (trin 5.3) og overgangen sandsynligheder er indstillet til 0,05 med undtagelse af sandsynligheder til ophold i den samme stat, som er valgt sådan at sandsynligheden for at forlade en bestemt stat er normaliseret til enhed.
En sandsynlighed profilering metode bruges til at give konfidensintervaller (CIs) for alle overgang priser21,22, der tjener som meningsfuld estimater for deres usikkerhed. For at beregne rammerne af CI for en bestemt frekvens, er overgang sandsynligheden af interesse fastgjort til en anden værdi end MLE. Dette giver test model λ’. En risiko ratio (LR) test sandsynligheden for 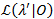 given datasæt 0 er udført i henhold til:
given datasæt 0 er udført i henhold til:
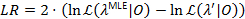
95% konfidensinterval bundet for parameteren er nået, når LR overstiger 3.841, 95% fraktil af en x2-fordelingen med en grad af frihed22,23.
Kraften i metoden er påvist ved hjælp af Hsp90. Dette rigeligt protein der findes i bakterier og eukaryoter og er en del af cellulært stress reaktion28. Det er en lovende drug target i cancer behandling29. Hsp90 er en homodimer med et nukleotid bindende lomme i hver underenhed30N-terminale domæne. Det kan gennemgå overgange mellem mindst to globalt forskellige konformationer, et lukket og et N-terminale åben, V-formet kropsbygning19,31,32. Dimerisk karakter rejser direkte spørgsmålet om samspillet mellem de to nukleotid-bindingssteder i Hsp90.
I følgende giver vi en trinvis protokol til dataopsamling og analyse af en trefarvet smFRET eksperiment på gær Hsp90 og nukleotid. Strukturafhængigt bindingen af fluorescently mærket AMP-PNP (AMP-PNP *, en ikke-hydrolyserbar analog af ATP) er analyseret. Anvendelse af den beskrevne procedure tillader undersøgelse af nukleotid bindende og samtidig Hsp90 konformationelle ændringerne og afslører dermed cooperativity mellem de to nukleotid bindende lommer af Hsp90.
