Ipossia, cioè, dissolto le concentrazioni ossigeno [] sufficientemente basso per influenzare negativamente biologico e processi ecologici ma spesso funzionalmente definita come [DO] < 2 mg / L1e anossia (funzionalmente definita come [DO] 0,0-0,2 mg/L) si verificano più frequentemente e gravemente in acque costiere del mondo, estuari e il profondo oceano2,3 e sono spesso aggravate dalla crescente eutrofizzazione4,5. Con un crescente estensione areale di ipossia e anossia, macrofauna sono influenzati negativamente e perdere la misura di habitat e qualità dell’habitat. Il cambiamento climatico è preveduto per peggiorare l’ipossia e anossia6.
In molti estuari stratificati, arricchita di sostanze nutritive come la Chesapeake Bay, Stati Uniti d’America, stagionalmente persistente ipossia può prevalere e può verificarsi anno dopo anno2. Inoltre, diel-ciclismo di ipossia è frequente negli estuari come la baia di Chesapeake e altri luoghi e si verifica tardi durante la notte o le prime ore del mattino in estate7,8.
Maggior parte degli studi si sono concentrati sull’effetto dell’esposizione continua di organismi a bassa [DO] e sulla loro tolleranza all’ipossia e anossia9,10,11,12,13,14 . Inoltre, gli studi hanno esaminato il cambiamento su larga scala nella composizione delle distribuzioni, abbondanze e specie specie in risposta a estesa [DO] basso4,15. Spesso specie che sono molto sensibili a bassa [], morire nelle masse,16 spostando le restanti specie di una fauna più giovane, più piccolo di dimensioni, di breve durata come, ad esempio, trovate sulla mensola Louisiana-Texas ecosistema4.
Cambiamenti comportamentali in genere precedono comunità crollo17 e gli studi hanno riferito sulle risposte comportamentali degli organismi a estesa basso [DO]4,16,17,18,19 ,20,21,22,23,24,25. Questi studi, tuttavia, non si concentrano sulle risposte degli organismi alle esposizioni diel-ciclismo di ipossia e la natura fluttuante di [DO] disponibilità negli estuari.
Diel-ciclismo ipossia negli estuari superficiali ha ricevuto crescente consapevolezza come studi monitorare [DO] più frequentemente nel corso dei giorni con sonde in estuari16,26. Acqua può rimanere ipossica per ore alla fine della notte o le prime ore del mattino in estate quando non c’è nessuna fotosintesi di generazione di ossigeno durante la notte, ma che consumano ossigeno alta respirazione aerobica7,16. Si è inoltre constatato che le maree influenzato il diel ciclismo condizioni basso con più estremi minimi osservati quando bassa marea ha coinciso con la fine della notte27. Solo dopo diverse ore di ipossia [DO] tornare a normoxia7,16,28 nel ciclo giornaliero.
Per determinare la risposta comportamentale di c. virginica diel-ciclismo ipossia e pH abbiamo monitorato l’apertura e la chiusura delle valvole di ostriche esposte al laboratorio indotto diel ciclica e ciclismo di [DO] pH. Gape risposte di bivalvi sono state utilizzate per rilevare condizioni ambientali avverse. Chiusure di valvola di molluschi bivalvi in risposta a contaminanti29,30,31, alghe tossiche32,33,34, l’inquinamento termico35,36 , 37,38,39,40, alimentazione regime39,41, emersione37,42, fotoperiodo43 diminuire la quantità di cibo , 44, pH45,46e combinato pH e ossigeno disciolto47 sono stati misurati. Gape tecniche hanno, ad esempio, incluso osservazioni dirette48,49,13, misurazioni continue utilizzando interruttori reed e magneti (Dreissena monitor)50o sensori in fibra ottica 51 che richiedono acqua limpida. Inoltre, sensori di Hall forza magneti e campi magnetici sono stati utilizzati per studiare la cozza gape angolo52,53,54,55e un sistema di induzione elettromagnetica ad alta frequenza che può misurare la distanza varia tra due bobine elettriche che sono incollati sulle valvole è stato utilizzato56,57,58,59. Una sorgente di alta tensione è necessaria per il sistema di induzione elettromagnetica e potenza deve essere consegnato ad entrambi i lati del guscio52. Questo sistema è anche disponibile nel commercio come il “MOSSELMONITOR” (http://mosselmonitor.nl/).
Un budget di ricerca stretto, abbiamo costruito un monitor economico strain-gauge (SGM) per misurare continuamente gape ostrica sopra indotta da laboratorio diel ciclismo di [DO] e pH, in condizioni di scarsa visibilità. Il nostro sistema è anche molto più semplice rispetto ai sistemi concorrenti, consentendo molti animali da instrumentare durante un esperimento. Abbiamo voluto determinare le risposte comportamentali di c. virginica diel ciclismo grave ([] = 0,6 mg/L) ipossia con pH di controllo (pH = 7.8) e ciclismo pH (pH = 7,8-7.0), rispettivamente e le risposte di gape al delicato ([] = 1,7 mg/L) ipossia. Inoltre, abbiamo voluto determinare se le ostriche sono in grado di rispondere rapidamente ai cambiamenti in [] sopra il diel ciclo e come reagire quando normoxia ritorna dopo un evento hypoxic. Forse le ostriche sono adattate all’ambiente rapidamente fluttuante che si trova in molti estuari16,27 dove vivono. Mentre più complessa valvola gape monitor sono disponibili, la SGM offre una tecnica poco costoso che consente misurazioni continue della valvola gape nelle acque anche in condizioni di scarsa visibilità.
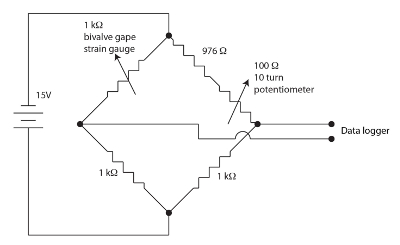
Figura 1. Ponte di Wheatstone per l’apparecchiatura di gape valvola. Clicca qui per visualizzare una versione più grande di questa figura.
I sensori dell’estensimetro utilizzati per il monitoraggio gape bivalvi sono film resistivo in un modello di meandro su un supporto di polyimide. Piccole quantità di sforzo modulano la resistenza del sensore. I bivalvi si flette l’estensimetro quando esso gapes causando un cambiamento nella resistenza del sensore. Abbiamo impiegato un ponte di Wheatstone Annulla, equilibrato, per ogni canale bivalvi come mostrato nella Figura 1 per misurare la variazione nella resistenza del sensore. Il ponte di Wheatstone è annullato dal potenziometro che permette un guadagno abbastanza alto essere impiegato dal datalogger. Un ponte di Wheatstone è un metodo standard per misurare con precisione una resistenza sconosciuta utilizzando un rapporto di un noto standard di resistenza e un voltmetro. La storia di questa antichissima tecnica è discussa in Ekelof (2001)60. Abbiamo integrato 12 canali, ognuno con il proprio ponte di Wheatstone e Annulla potenziometro, Strain Gauge Monitor (SGM).
