Hypoxi, dvs upplöst syre koncentrationer [göra] tillräckligt lågt för att negativt påverka biologiska och ekologiska processer men ofta funktionellt definierade som [göra] < 2 mg / L1och anoxia (funktionellt definierade som [DO] 0,0-0,2 mg/L) sker oftare och hårdare i världens kustvatten, flodmynningar och den djupa hav2,3 och förvärras ofta av ökande eutrofiering4,5. Med en ökande areal grad av hypoxi och anoxi, makrofauna påverkas negativt och förlora livsmiljö omfattning och livsmiljö kvalitet. Klimatförändringarna förutspås för att förvärra hypoxi och anoxi6.
I många stratifierat, näringsämne-berikad flodmynningar som Chesapeake Bay, USA, säsongsmässigt ihållande hypoxi kan segra och kan uppstå efter år2. Dessutom diel-cykling av hypoxi är frekventerar i flodmynningar som de Chesapeake Bay och andra platser och uppstår sent under natten eller tidigt på morgonen i sommar7,8.
De flesta studier har fokuserat på effekten av kontinuerlig exponering av organismer till låg [DO] och deras tolerans till hypoxi och anoxi9,10,11,12,13,14 . Studier har dessutom tittat på storskalig förändring i artsammansättningen distributioner, sammansättning och arter som svar på utökade låg [DO]4,15. Ofta arter som är mycket känsliga för lågt [göra], dö i massor,16 skiftande övriga arter till en yngre, mindre och medelstora, kortlivade fauna som till exempel, hittade på Louisiana-Texas hylla ekosystem4.
Beteendeförändringar vanligtvis föregår gemenskapens kollaps17 och studier har rapporterat på beteendemässiga Svaren av organismer till utökade låg [DO]4,16,17,18,19 ,20,21,22,23,24,25. Dessa studier, dock fokuserar inte på svaren från organismer till diel-cykling exponeringar av hypoxi och fluktuerande natur [DO] tillgänglighet i flodmynningar.
Diel-cykling hypoxi i grunt flodmynningar har fått ökande medvetenhet som studier övervaka [göra] mer ofta under loppet av dagar med Kerstin i flodmynningar16,26. Vatten kan förbli hypoxisk för timmar i slutet av natten eller tidigt på morgonen på sommaren när det finns ingen genererar syre fotosyntesen under natten men hög syreförbrukande aerob respiration7,16. Det konstaterades också att tidvattnet påverkas diel cykling låg villkor med de mest extrema minima observerade när lågvatten sammanföll med avsluta av natten27. Först efter flera timmar av hypoxi [gör] komma tillbaka till normoxia7,16,28 i dagligt cykeln.
Att bestämma C. virginica beteendemässiga svar på diel-cykling hypoxi och pH vi övervakade öppning och stängning av ventiler av ostron utsätts för laboratoriet inducerad diel cykling [Do] och cykliska pH. Gapa Svaren av musslor har använts för att upptäcka skadliga miljöförhållanden. Ventil nedläggningar av musslor som svar på föroreningar29,30,31, giftiga alger32,33,34, termisk förorening35,36 , 37, mat kvantitet minska38,39,40, utfodring regimen39,41, emersion37,42, fotoperiod43 , 44, pH45,46, och kombinerade pH och löst syre47 har mätts. Gapa tekniker har, till exempel ingår direkta observationer48,49,13, kontinuerliga mätningar med reed switchar och magneter (Dreissena monitor)50eller fiberoptiska sensorer 51 som kräver klart vatten. Dessutom har magnet och magnetfält styrka hallgivare använts för att studera mussla gapa vinkel52,53,54,55och ett högfrekvent elektromagnetisk induktion system att kan mäta varierande avståndet mellan två elektriska spolar som är limmade på ventilerna har varit används56,57,58,59. En hög spänningskälla krävs för elektromagnetisk induktion system och makt måste levereras till båda sidor av shell52. Detta system är också kommersiellt tillgängliga som den ”MOSSELMONITOR” (http://mosselmonitor.nl/).
På en tight forskningsbudget, konstruerade vi en billig töjningsmätaren monitor (SGM) att kontinuerligt mäta oyster gapa över laboratorium-inducerad diel cykling [Do] och pH, låga siktvärden villkor. Vårt system är också mycket enklare än konkurrerande system, vilket möjliggör många djur att vara instrumenterad under ett experiment. Vi ville bestämma de beteendemässiga svar på C. virginica diel cykling svår ([göra] = 0,6 mg/L) hypoxi med kontroll pH (pH = 7,8) och cykling pH (pH = 7,8-7.0), respektive, och gapa Svaren till mild ([göra] = 1,7 mg/L) hypoxi. Dessutom ville vi avgöra om ostron är kunna svara snabbt på förändringar i [göra] över diel cykel och hur de reagerar när normoxia återgår efter en hypoxisk händelse. Kanske är ostron optimalt anpassade till snabbt fluktuerande miljö som finns i många flodmynningar16,27 där de lever. Medan mer komplexa ventil gapa bildskärmar finns, erbjuder SGM en billig teknik som tillåter kontinuerliga mätningar av ventilen gapa i vatten även i låga siktvärden förhållanden.
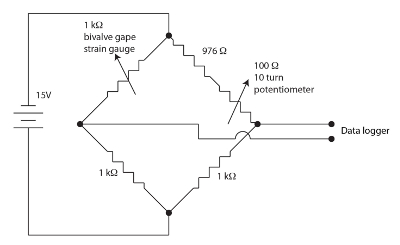
Figur 1. Wheatstone bridge för ventil gapa apparatur. Klicka här för att se en större version av denna siffra.
Anstränga mätinstrumentet sensorerna används för övervakning av tvåskaliga gapa är resistiv filmer i en meander mönster på en polyimid bakningen. Små mängder stam modulera motståndet av sensorn. Tvåskaligt flexar stam nivåglaset när det gapar orsakar en förändring i sensorns motstånd. Vi anställt en ogyltig, balanserad, Wheatstone-brygga för varje tvåskaliga kanal som visas i figur 1 att mäta förändringen i temperatursensor motståndet. Wheatstone överbrygga är nulled av potentiometern tillåter en ganska hög vinst att vara anställd av en datalogger. En Wheatstone bridge är en standardmetod för att exakt mäta en okänd motstånd med hjälp av ett förhållande till en känd motstånd som standard och en voltmeter. Historien om denna mycket gamla teknik diskuteras i Ekelof (2001)60. Vi integrerat 12 kanaler, alla med sin egen Wheatstone bridge och ogyltig potentiometer, töjningsmätaren Monitor (SGM) enheten.
