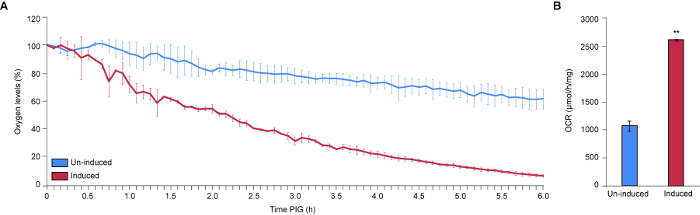
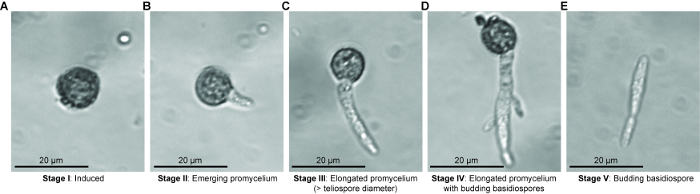
Basidiomycete biotrophic plant pathogens cause billions of dollars in crop losses annually. The vast majority of these pathogens produce teliospores that are integral to fungal dispersal and sexual reproduction. Gaining knowledge of the development and germination of teliospores is critical to understanding the spread of the devastating diseases caused by these fungi. In order to identify molecular changes at key control points we have devised a method to identify the timing of physiological shifts and another to isolate teliospores at distinct stages of germination. Seto et al. (unpublished) noted five stages of teliospore germination by light microscopy (Figure 2). In order to investigate physiological activation during Stage I and to assess respiration rate during germination, we used a Clark-type microrespirometer to precisely measure changes in oxygen consumption. Our sample data indicate that our method is precise and highly reproducible. Our findings confirm that germinating U. maydis teliospores exhibit a drastic increase in cellular respiration compared to un-induced dormant teliospores. For the first time, we have identified that U. maydis teliospores that have been induced to germinate exhibit a ~45 min delay in oxygen uptake. This suggests that U. maydis teliospores may require some time to process germination signals (e.g., the presence of sugars) before responding, that increased oxygen uptake is not among the very immediate responses to germination signals or that our assay was not sensitive enough to detect the minimal change in initial oxygen uptake.
Previous studies examining respiration rates of smut teliospores3 relied on a Warburg flask apparatus to measure oxygen levels manometrically5. Briefly, this method measures oxygen consumption and CO2 production by detecting changes in pressure in an enclosed flask through the direct observation of fluid level changes in the manometer arm. The experiments can be difficult to set up, and measurements can be imprecise. The apparatus must be attended throughout the period of measurement and, extensive calculations are required to estimate OCR. Our protocol makes use of technological advances, eliminating the requirement for the user to remain by the apparatus for the duration of the experiment, take measurements by eye, and use extensive mathematical formulas. Others have used early Clark-type respirometers to measure OCR of Neurospora crassa9 and Botryodpilodia theobromae10 spores, however, these early instruments permitted continuous measurements for a maximum of 20 min. This limitation would not have allowed the identification of the ~45 min delay in oxygen uptake we observed with the newer model respirometer. Our protocol has made data interpretation simpler, as the readout is the concentration of oxygen remaining in the chamber, which can be directly graphed without any calculations or data manipulation. In addition, it is possible to take continuous measurements (every 2 s) for an indefinite amount of time until available oxygen is completely depleted. This permits the identification of small changes in respiration over a long period of time. Therefore, we have improved upon earlier techniques and developed a simple, precise, and reproducible method to measure oxygen consumption of fungal spores. To our knowledge, this is the first study to use a modern Clark-type respirometer to study respiration of dormant versus germinating teliospores of smut fungi.
Despite the ease and simplicity of this protocol, optimization is required and there are biological realities that limited the analysis. First, appropriate sample sizes must be identified to achieve reasonable OCRs. Too much sample can lead to premature crashing of oxygen levels, and too little sample can result in the inability to observe meaningful changes in oxygen consumption. Second, it is imperative to allow the probe time to stabilize (~3 min) to provide accurate initial data. Lastly, it is important to supplement germination medium with antibacterial agents (e.g., streptomycin sulfate) in order to ensure bacterial contamination does not alter OCR readings. The biological limitations we faced were a low germination rate over the time course of measurement, (~1%) as determined by observing visual morphological changes. Determining spore viability would allow this rate determination to be converted to a rate per spore number and isolating teliospores with higher rates of germination would lead to higher OCRs. The asynchronous germination of U. maydis teliospores11 is a reality that must be accounted for and may have contributed to an inability to detect oxygen consumption earlier in germination.
In order to improve the accuracy and precision of measuring changes in teliospore respiration, future adaptations to this method could include measuring OCR on a single cell-basis. Micromanipulation techniques could be used to isolate a single teliospore, which can then be induced to germinate, and its respiration rate can be monitored. This could improve resolution, providing information regarding the OCR during the dormancy-germination shift per teliospore, rather than per mg of teliospores. In addition, this would solve the confounding issue of asynchronous germination.
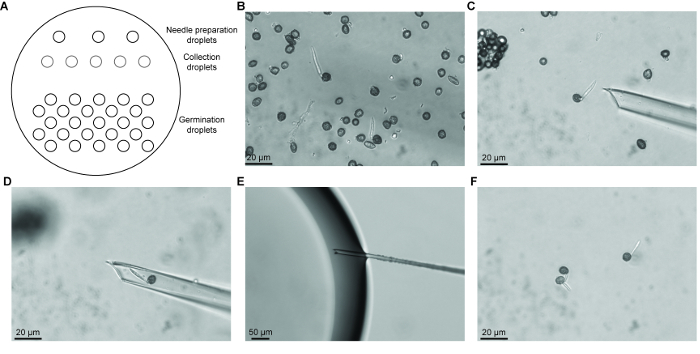
For later stages of germination, we developed a micromanipulation method to isolate teliospores at common stages of germination. This allowed the creation of relatively synchronous teliospore populations for analysis. Various methods for isolating single microorganisms have been described and have been improved upon over the years12. These methods include the dilution of spore suspensions to obtain single microorganisms, semi-mechanical methods with the use of microcapillaries to obtain spores that are transferred to medium for culturing, and mechanical methods which use micromanipulators. Previous methods that we used to obtain teliospores at the same stage of germination include counterflow centrifugal elutriation and filtering germinating teliospores through a nylon membrane with a specific pore size. Using these methods allowed us to enrich for germinating teliospores, however, our samples still contained teliospores in various stages of germination13. Current technology for micromanipulation of single microorganisms has improved with the introduction of higher magnification and instruments for fine control of capillary needles, aspiration, and transfer of microorganisms. Previous micromanipulation techniques have focused on isolating single cells for culturing or for use in single cell PCR applications14. The use of micromanipulators to isolate single fungal spores has not previously been established. A previous method for isolating single fungal spores involved the use of fine forceps or needles to pick small pieces of solid medium containing germinating spores15. Micromanipulation with the use of micromanipulators is widely used in yeast studies where clusters of ascospores can be separated following sporulation in culture on agar medium for meiotic genetic analysis16. We have developed a method which combines the micromanipulation technique for bacterial cells14 and in vitro fertilization methods for isolating germinating teliospores. We have shown that hundreds of common germination stage teliospores can be obtained with this technique. These samples can be used for downstream expression studies using techniques such as RT-qPCR or RNA-seq. Obtaining a population of teliospores in which germination is synchronized permits the analysis of specific changes in gene expression that occurs during early, mid and later stages of teliospore germination.
Microdissection of stage specific germinating teliospores may require experience in set up and recognizing the different stages of germination, however, this experience can be obtained quickly through practice. There are several steps that must be followed for successful microdissection followed by RNA isolation. First, germination medium must be supplemented with antibiotics (e.g., streptomycin sulfate) to suppress the growth of bacterial contamination when germination is initiated as well as during collection of germinating teliospores. Second, it is important to use a stabilization solution to stabilize and protect RNA for isolation. The RNA stabilization solution also prevents collected teliospores from progressing to the next germination stage while collecting additional teliospores. Thirdly, it is important to remove the mineral oil once the collection droplet has been recovered to ensure successful RNA extraction. Lastly, we have noticed some loss of RNA quality if isolated teliospores are stored in RNA stabilization solution for an extended period of time; therefore, it is recommended that RNA is isolated immediately following collection of germination stage specific teliospores. A limitation of the method is that the Stage I teliospores collected could contain dormant, dead, and induced to germinate teliospores as these three stages are morphologically indistinguishable. In addition, when collecting Stage III teliospores, a mixture of teliospores in meiosis I or meiosis II could be obtained. One way to aid in distinguishing between truly dormant and dead teliospores could be to determine the viability of the sample. A method for assessing fungal spore viability using live/dead cell viability assays may be able to assess percentage of viable teliospores from which more informative germination rates could be determined17. In addition, nucleus staining with DAPI, for example, could be used to visualize the events of meiosis that are occurring during Stage III and the transition to Stage IV in order to further characterize teliospores morphologically at Stage III. This would aid in the collection of teliospores in only one stage of germination when using our microdissection method.
In conclusion, we have developed a simple, precise and reproducible method of measuring the changes in cellular respiration that occur during the dormancy-germination shift of Ustilago maydis teliospores. In addition, we have developed a method for collecting specific stages of germinating teliospores that could be used for downstream applications, such as RNA-seq. Our methods can be adapted to accommodate various cell-types and species. We anticipate that improvements to our techniques will facilitate the detection of respiratory changes on a single spore level as well as further defining the events that are occurring in the later stages of germination.