单核细胞, 中性粒细胞和树突状干细胞 (DCs) 是由造血祖, 主要是在骨髓中, 由一个称为 myelopoiesis 的过程中产生的髓体细胞。常见的髓样祖细胞 (中医) 有可能产生髓体细胞, 以及巨和红细胞, 而不是淋巴细胞。粒细胞-单核细胞祖 (GMPs) 是由中医衍生而成的, 其产生的是颗粒和单核细胞, 但失去了巨和红细胞电位。单核细胞和经典和浆 DCs (招揽/pDCs) 也被认为是由一般的祖细胞-DC 祖 (mdp 以) 产生的, 这是由中医生产的. 世系势的逐步限制最终导致沿袭前身: 粒细胞祖, 单核细胞祖, 树突状体细胞祖 (图 1)。
魏斯曼和同事报告说, 中医是在林 c-试剂盒+ Sca-1– (LKS) CD34+ FcγR 的小鼠骨髓部分, 而 GMPs 包含在 LKS CD34 + FcγR嗨分数1。然而, 这些 “CMP” 和 “GMP” 分数是非常异构的。例如, “GMP” 分数也包含血统承诺粒细胞祖和单核细胞祖1,2。mdp 以分别被报告为 CX3CR1+ Flt3+ CD115+祖细胞也表达 CD34 和 FcγR3,4。mdp 以产生了 cDC/pDC 产生的共同 DC 祖 (CDPs), 据报道, 它表示较低水平的 c 套件 (CD117), 并没有包括在 LKS 分数5.
以前假定单核细胞是通过单一途径产生的 (CMP-GMP-MDP 单核细胞)。与这个模型相一致的是, 由 GMPs (命名的单核细胞祖, MPs)2和 mdp 以 (命名的共同单核细胞祖, cMoPs) 产生的单核细胞的祖细胞,6在共享表面标记表达式的基础上似乎是同一单元格.然而, 我们最近证明单核细胞是由 GMPs 和 mdp 以独立产生的, 并且能够通过单细胞 RNA 测序7区分 MPs 和 cMoPs。
我们最近修改了魏斯曼 “CMP” 和 “GMP” 浇口战略, 以确定6脂蛋白亚组分的 C57BL/6J 小鼠骨髓含有不同的 oligopotent 和血统承诺髓祖亚群。我们首次报告说, Ly6C 和 CD115 染色允许隔离 oligopotent GMPs, 以及粒细胞祖 (GPs) 和单核细胞祖 (我们目前无法分离的 MPs 和 cMoPs) 从 “GMP” 分数2 (LKS–CD34+ FcγRhi门;图 1)。我们随后证明, mdp 以主要见于 “CMP” 分数 (LKS– CD34+ FcγRlo门), 其中还包含 Flt3+ CD115lo和 Flt3–子集7 (图1).CMP-Flt3+ CD115 的 GMPs 和 mdp 以在收养转移时产生。CMP-Flt3 子集包含的祖细胞似乎是中间体之间的 CMP-Flt3+ CD115 和 GMPs。与 mdp 以不同, CMP-Flt3+ CD115 和 CMP-Flt3 分数也具有巨和红细胞电位。
然而, 必须指出的是, 目前尚不清楚 “CMP” 分数是否包含真正 oligopotent 的祖细胞 (例如, CMP-Flt3+ CD115 中的单个单元, 它拥有中性粒细胞,单核细胞、DC、巨和红细胞电位), 或二者择一地, 构成了一个具有更限制性血统潜能的祖先的混合物。菌落形成检测 (纤维素培养) 揭示了粒细胞 (中性粒细胞), 红细胞, 单核和巨电位 (GEMM 细胞) 在 “CMP”, CMP-Flt3+ CD115lo和 CMP-Flt3–分数1 ,7、但不允许对直流电位进行评估。与此相反, 菌落形成检测表明, 在 “GMP” 分数1,2, oligopotent GMPs (祖细胞和单核细胞潜能) 的存在, 这是支持的最近单细胞transcriptomic 分析8。然而, 目前还不知道这些 oligopotent GMPs 是否也产生其他粒细胞 (嗜酸性粒、嗜碱性细胞和肥大的干细胞)。
在这些研究的基础上, 我们现在展示了7表面标记 (c 套件, Sca-1, CD34, FcγR, Flt3, Ly6C 和 CD115) 可以用来识别和分离这6个子集的 oligopotent 和血统承诺的髓细胞祖细胞。这里描述的协议可以应用于体外培养试验 (纤维素或液体培养), 在小鼠体内移植实验, 分子分析 (块状和单细胞 RNA 测序, 西方印迹, 等)。
该协议由3个阶段组成: 1) 制备骨髓细胞单细胞悬浮液, 2) 对造血祖 (磁性活化细胞分选) 和 3) 进行富集鉴定, 并根据需要隔离祖子集, 通过流动细胞术 (根据需要使用分析仪或分拣机)。第一步是从安乐死小鼠的股骨和胫骨中分离骨髓细胞, 类似于先前描述的9项协议。接下来, 通过对红细胞、中性粒细胞、单核细胞、淋巴细胞等细胞表面标志物的抗体, 对茎和祖细胞进行浓缩, 以耗尽分化的细胞。这不是强制性的, 但强烈建议优化检测的祖子集, 并减少所需的抗体数量的祖鉴定和流式细胞术所需的时间。下面的沿袭损耗协议描述了磁活化细胞分类 (mac) 使用鼠标谱系细胞耗尽套件 (其中包含生物素化抗体对 CD5, CD45R (B220), CD11b, Gr-1 (Ly6G/C), 7-4 和 Ter-119, 外加抗生物素微珠) 和自动磁选机。最后一步是流式细胞术对祖子集的识别 (和排序, 如果需要的话)。下面描述的抗体面板 (另见表 1) 已设计用于流式细胞仪 (分析仪或分拣机) 与4激光器 (405 nm, 488 nm, 561 nm, 640 nm)。
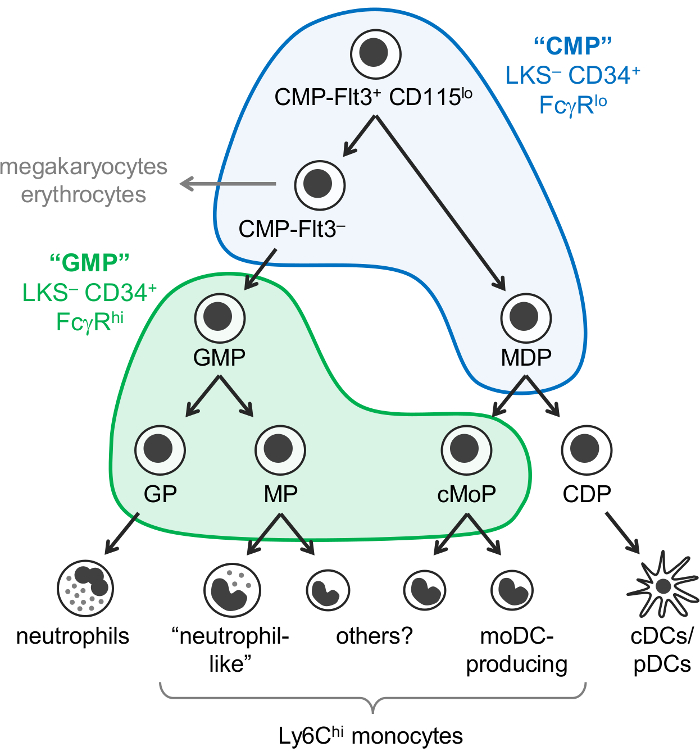
图 1: 中性粒细胞、单核细胞、DC 祖和分化通路.最近修订的 myelopoiesis7模型被说明, 魏斯曼门为 “中医” (蓝色) 和 “GMPs” (绿色)1覆盖。这一数字已从亚涅斯等20177修改。请单击此处查看此图的较大版本.
