Probe recovery
The mean recovery (i.e., the mean amino acid content in the perfusate as a percentage of the content in an equal volume of the vial solution) was 15.49 ± 0.42% at a flow rate of 2 μL/min and 6.32 ± 0.64 at 3 μL/min for glutamate and 14.89 ± 0.36% at a flow rate of 2 μL/min and 10.13 ± 0.51 at 3 μL/min for aspartate when using the cuprophane membrane probe. If using the polyacrylonitrile membrane probe, the mean recovery was 13.67 ± 0.42% at a flow rate of 2 μL/min and 6.55 ± 1.07 at 3 μL/min for glutamate and 14.29 ± 0.62% at a flow rate of 2 μL/min and 8.49 ± 0.15 at 3 μL/min for aspartate (Figure 4A-4B). As it can be clearly seen in Figure 4A, the slower flow rate (2 μL/min) enhances the dialyzing performance of both probes. For the following experiments the cuprophane membrane endowed probe perfused at a flow rate 2 μL/min was chosen, because its mean recovery was higher (even if not significantly) at this flow rate for both analytes and because of experimental continuity (these probes were used for analyzing GABA in precedence1).
Seizures development and progression of the disease after status epilepticus
The behavioral and EEG monitoring of seizures, their evaluation, was done in all the animals employed in this study to confirm the development and progression of TLE disease in these.
The robust convulsive SE, that was interrupted after 3 h using diazepam, occurred 25±5 min after the pilocarpine injection. Then, all the animals entered a latency state in which they were apparently well and they were continuously video-EEG monitored in order to verify that no spontaneous seizures occurred in the first 9 days or to identify the first spontaneous seizure, respectively for the latency and the first seizure group. The first spontaneous seizure occurred 11.3 ± 0.6 days after SE (mean ± SEM, n=21). Thereafter, seizures occurred in clusters, and aggravated in time. In late chronic phase (days 55-62 after SE) the epileptic rats experienced 3.3±1.2 (mean ± SEM, n=12) generalized seizures daily. There was a clear progression of the disease. Many, but not all EEG seizures, corresponded with behavioral seizure activity. Figure 5B shows the recorded paroxysmal epileptiform activity that was observed about 500 ms before and during behavioral seizures. Figure 5A shows control traces in non- epileptic rats.
Representative basal values of amino acids found in microdialysis perfusate and potassium stimulated release of glutamate
Basal glutamate concentration found in chronic epileptic rats (0.87 ± 0.06 µM) was significantly higher than in control animals (0.59 ± 0.03 µM; p < 0.05 vs. controls; one-way ANOVA and post hoc Dunnett's test). There was no statistically significant difference between chronic epileptic (0.31 ± 0.04 µM) and non-epileptic animals (0.30 ± 0.05 µM) in basal or high K+ evoked aspartate concentrations. See the original article for details2. The reported basal levels of glutamate in control rats are in line with those found by others in similar studies (i.e., about 0.75 µM when using a 2 µL/min flow and membranes of 2 mm effective length)46,47,48,49,50,51,52,53. However, many different factors can influence the results of microdialysis, for example the effective length of the probe and the membrane cut off.
High K+-evoked an additional release of glutamate for about 30 min in control rats and for about 60 min in chronic epileptic rats (Figure 6). See the original article for details2. As can be seen from the depicted time course, the 10 min time resolution of microdialysis was sufficient to capture the variances in glutamate release found in both groups of animals.
HPLC calibration and limits
The data were calculated based on calibration curves obtained with standard solutions of glutamate and aspartate and the internal standard L-homoserine. The concentration of the neurotransmitters glutamate and aspartate in the perfusates was expressed in absolute values (µmol/L). Each calibration plot was constructed by analysis of solutions of glutamate and aspartate at four concentration levels (five replicates at each level).Regression coefficients were calculated for calibration plots: y = kx + q, where x was the concentration ratio of aspartate or glutamate to L-homoserine (IS) and y was the corresponding peak-area ratio of aspartate or glutamate to L- homoserine (IS). The coefficient of determination (r2) was calculated. The applicability of HPLC method was within the limits; the lower limit of quantification was determined as the lowest concentration in the standard calibration curve and the upper limit of quantification as the highest used concentration of amino acid analytes for calibration, respectively. Limit of detection (LOD) was also calculated. Some of these values are delineated in Table 1. A model chromatogram of blank sample, standards sample and collected dialysis sample obtained with above described method are shown in Figure 7.
Probe localization
Microdialysis probe and recording electrode were implanted into the right ventral hippocampus and their correct placement was verified. Only those animals where the implantation was maximally in 500 μm distant from stabilized coordinates (see Figure 8) were included in analysis.

Figure 1. Step-by-step preparation of the device to be implanted. (A) 3-channel electrode with 10 cm long grounding electrode in its protective sleeve on the left and guide cannula for microdialysis on the right needed to assemble the device. (B). The bare electrode and guide cannula in detail. The first step is to remove (C) and insert (D) the metal guide cannula from and into its plastic dummy few times to ease its release once implanted into the animal's head. The second step is to bend two times the twisted registering electrode to be aligned to dialysis guide cannula (E, F). (G) The electrode tip should be cut to be 0.5 mm longer than the tip of the metal guide cannula. (H) Check for the precision of the cut using the digital caliper. Subsequently, about 1 mm long silicon circlet should be used to fix the alignment of electrode to guide cannula foot (I). (J) The photograph showing how to ring the electrode and guide cannula shaft. The final step is to put a drop of resin or glue onto the guide cannula pedestal fixing the silicon circlet to it (K). (L) Assembled device ready to be sterilized. Please click here to view a larger version of this figure.

Figure 2. Photographs of different types of devices for microdialysis-EEG in rats used (A) and the photograph of the probe clip holder (B) used to implant these devices. (A) The guide cannula (in green) is replaced by a microdialysis cannula typically 24 h before the experiment. The electrical connector of the device (first left was used for the recordings described in this manuscript) permits the attachment of wires that conduct electrical signal to amplifier and data collection equipment. The device is surgically attached to the skull of anesthetized rats and recordings may be obtained later without causing pain or discomfort in freely behaving rats. Please click here to view a larger version of this figure.
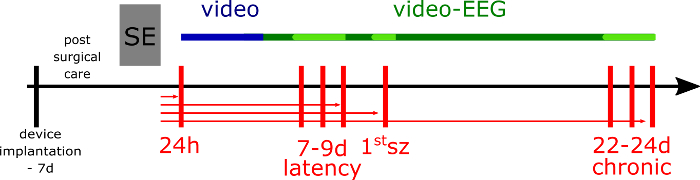
Figure 3. Experimental design. The week before status epilepticus (SE) induction, the rats are implanted with the device. SE is induced by pilocarpine and animals (if not dialyzed and killed at 24 h after SE; the rats from acute group) are video monitored for 5 days (blue line), then video-EEG monitored to assess the seizure frequency and duration in their home cages (green line). For the microdialysis experiment, the epileptic and respective non-epileptic control rats are transferred to another EEG set up equipped with cylinder cages in 24 h before the microdialysis session and still video-EEG monitored (light green line). The vertical red lines represent the dialysis sessions at different time-points of epileptic disease development. The horizontal red lines represent the different groups of epileptic animals (and respective non-epileptic animals), where the arrow indicates the last day of the microdialysis and the day of animal's death. Please click here to view a larger version of this figure.
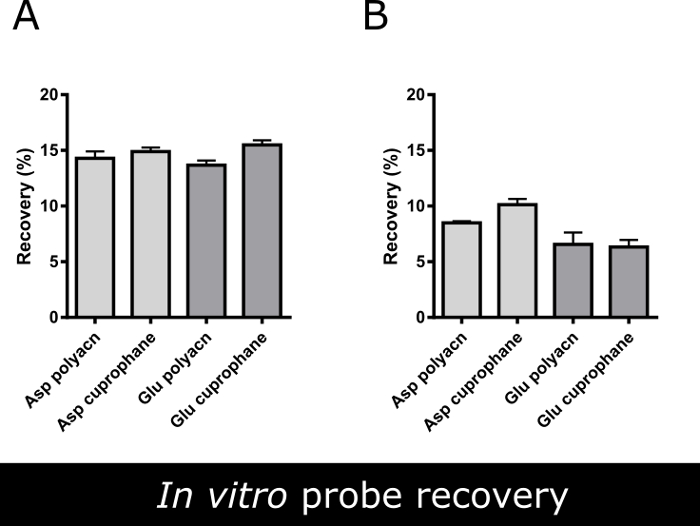
Figure 4. In vitro recovery of two dialysis probes. Mean in vitro recovery (%) of aspartate and glutamate using two different commercially available microdialysis probes (both endowed with 1 mm long dialyzing membrane) at (A) 2 μL/min and (B) 3 µL/min flow rate. Data are the mean ± SEM of 3 independent experiments run in triplicates. There are not statistically significant differences between the efficiency of various probes (Student's unpaired t-test, p<0.05).Using a flow rate 2 μL/min the glutamate recovery increased about 5% compared to 3 µL/min flow rate, thus the slower flow rate was used for microdialysis experiments. Please click here to view a larger version of this figure.
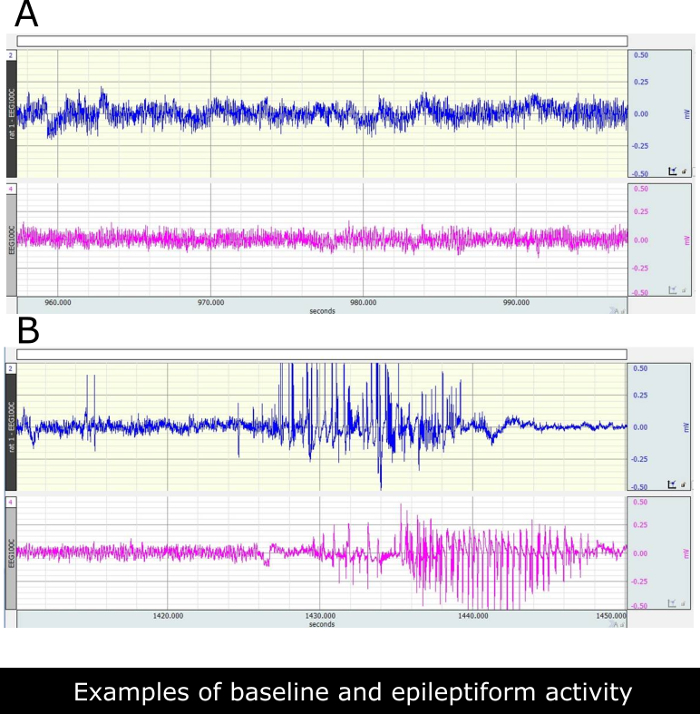
Figure 5. Illustrative EEG recordings from ventral hippocampus of paraoxystic activities as can be seen at chronic phase in control and epileptic rats. (A) Two representative traces recorded in two saline treated non-epileptic rats. (B) Traces recorded in two epileptic rats. Epileptiform discharges correspond with class 3 behavioral seizures in these rats. Please click here to view a larger version of this figure.
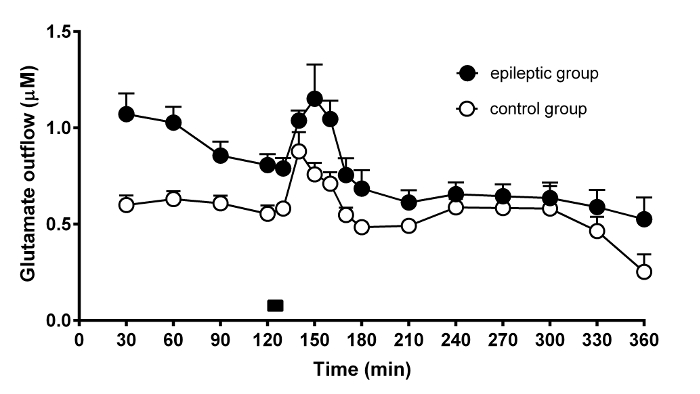
Figure 6. Time-course of the effect of potassium stimulation on glutamate release from the rat hippocampus. Representative result of the microdialysis experiment performed in 6 control (open circles) and 6 chronic epileptic rats (black circles). The graph shows the temporal changes of dialysate glutamate concentration in the course of microdialysis experiments and during high 100 mM K+ stimulation. The time of high K+ stimulus (10 min) is indicated by the black bar on bottom of the graph. The data are the means ± SEM of 6 animals per group. Please click here to view a larger version of this figure.
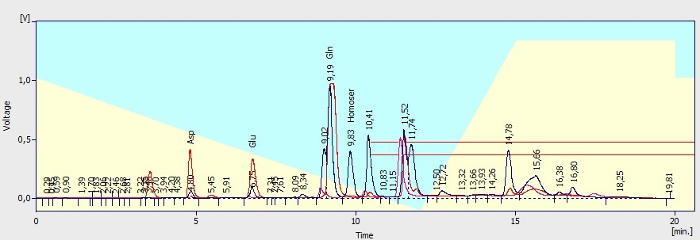
Figure 7. Illustration of chromatograms. Known peaks are labeled. Pink trace: chromatogram of Ringer's solution without intentionally added amines after OPA/5-ME derivatization (blank sample). Blue trace: chromatogram of dialysate sample after derivatization showing the peaks of amino acids: aspartate (tR 4.80 min), glutamate (tR 6.75 min) and glutamine (tR 9.19 min) and the peak of IS L-homoserine (2.5 µM, retention time, tR 9.83 min). Red trace: Chromatogram of standard of aspartate (2.5 µM) and glutamate (2.5 µM) in Ringer's solution. Azure and yellow background of the picture stands for mobile phase A (azure) and mobile phase B (yellow) portion used for analytes elution. A red rectangle indicated area (tR 10.41 min and further) shows the peaks of unknown substances and OPA degradation products. All injection volumes were 20 µL. The derivatives were separated at a flow rate of 0.8 mL/min. Please click here to view a larger version of this figure.
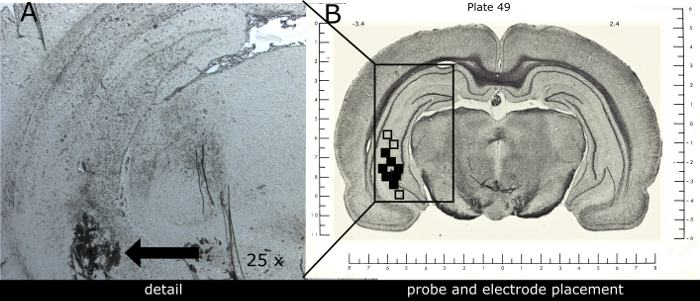
Figure 8. Representative image of combined electrode-probe placement within the ventral hippocampus. (A) Photograph shows the scare left by the device tip in detail (black arrow). (B) Schematic illustration of the electrode-probe tip positions within the implanted ventral hippocampus of 12 rats. The solid squares (some overlapping) indicate correctly localized probe-electrode tips. Open squares indicate incorrectly localized probe-electrode tips in animals excluded from the study (n=3). Coronal brain slices containing probes and recording sites were processed after experiments for histological analysis. The numbers above the illustration show the distance from Bregma (according to Pellegrino et al. 1979 atlas of rat brain; nose bar + 5.0 mm, co-ordinates used: A -3.4 mm, L+5.4 mm; P + 7.5 mm from dura). Please click here to view a larger version of this figure.
| Analyte | c (μmol/l) | k | q | r2 | LOD (pmol/l) |
| Glutamate | 0.25-2.5 | 5.215 | 1043.79 | 0.999 | 19.4 |
| Aspartate | 0.25-2.5 | 2.258 | 1994.72 | 0.998 | 31.7 |
Table 1. Quantification characteristics of HPLC method used for amino acids determination. Concentration range of standards (c), slope (k), intercept (q), coefficient of determination (r2) and limit of detection (LOD) describing the calibration plots obtained with standard solutions of glutamate and aspartate (0.25, 0.5, 1.0 and 2.5 µM) and internal standard L-homoserine (2.5 µM) using the described HPLC method with spectrofluorometric detection.
