Two methods were employed: an ATP utilization assay and a live/dead staining assay. The results can either be combined or individually used for examining bacterial survival within aggregates after antibiotic treatment. The ATP utilization assay has been shown to measure accurately viable bacteria in S. aureus biofilms20,21. Here, MS11Opa+Pil+ strain was used to examine the role of GC aggregation in antibiotic susceptibility. Non-aggregated MS11Opa+Pil+, aggregated MS11Opa+Pil+, or aggregated and then disrupted by sonication MS11Opa+Pil+ were treated with serial dilutions of ceftriaxone and the ATP level measured (Figure 1A). In comparing the percent survival with and without antibiotic treatment, pre-aggregated GC had significantly higher survival than non-aggregated or aggregation-disrupted GC with equal at or above 0.015 µg/mL of ceftriaxone (MIC from agar dilution test (Table 4)). MS11Opa+Pil-, MS11ΔOpa or MS11ΔLgtE, which have the same agar dilution MIC (Table 4), but form smaller aggregates, was examined and compared to MS11Opa+Pil+ (Figure 1B). MS11Opa+Pil+, forming larger aggregates, had the higher ATP level with ceftriaxone treatment than the mutant strains.
Live/dead staining has been used in several biofilm/aggregation-related studies22,23.To determine the effect of aggregation, pre-aggregated MS11Opa+Pil+ was treated with or without ceftriaxone and imaged. This allows both visualization (which can be quantified) and the distribution of live and dead GC after antibiotic treatment (Figure 2A– left two panels). Dead bacteria (red) were largely located at the outer layers whereas live GC (green) were located mainly in the core of ceftriaxone treated aggregates. This procedure was performed with MS11Opa+Pil-, MS11ΔOpa or MS11ΔLgtE, to examine aggregation size and survival rate (Figure 2A). MS11Opa+Pil+ was shown to form the largest and MS11Opa+Pil- the smallest aggregates (Figure 2A,B). MS11Opa+Pil+ aggregates were still alive in the core layer whereas GC in the small loose aggregates of MS11ΔOpaPil+, MS11Opa+Pil-, and MS11ΔLgtEPil+ were dead (Figure 2A,C). Based on the size and survival, a correlation graph can be plotted to examine the relationship of aggregation size and antibiotic survival (Figure 2D).

Table 1: Recipe for 1 L of GCK Agar Plate.

Table 2: Recipe for 1 L of 100x Kellogg's supplement.
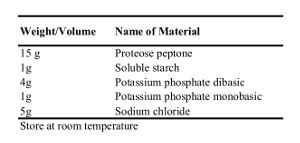
Table 3: Recipe for 1 L of GCP Bacterial Growth Media.

Table 4: Minimum inhibitory concentration of GC strains treated with ceftriaxone. MS11Opa+Pil+, MS11ΔOpa, MS11ΔLgtE, and MS11Opa+Pil- were grown and suspended in GCP. Agar dilution test was then performed with serial concentration of ceftriaxone from 0.0016 – 0.25 µg/mL.
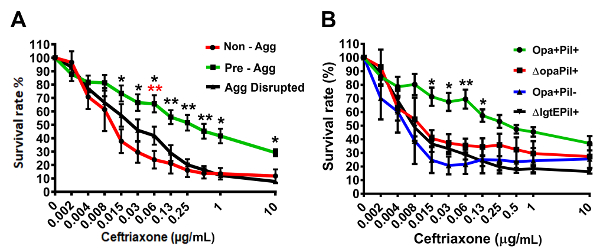
Figure 1: Representative data of survival rate of aggregated GC under ceftriaxone treatment by ATP-utilization assay. (A) Survival rate comparison of MS11Opa+Pil+ suspension without pre-aggregating, pre-aggregating for 6 h, or disrupting after pre-aggregating for 6 h. (B) Survival rate comparison of 6 h aggregated MS11Opa+Pil+ with MS11ΔOpaPil+, MS11Opa+Pil-, or MS11ΔLgtEPil+. Shown are the average values (±SD) obtained from three independent experiments. ***p < 0.001; **p < 0.01; *p < 0.05. This figure was previously published 24 and is used with permission. Please click here to view a larger version of this figure.
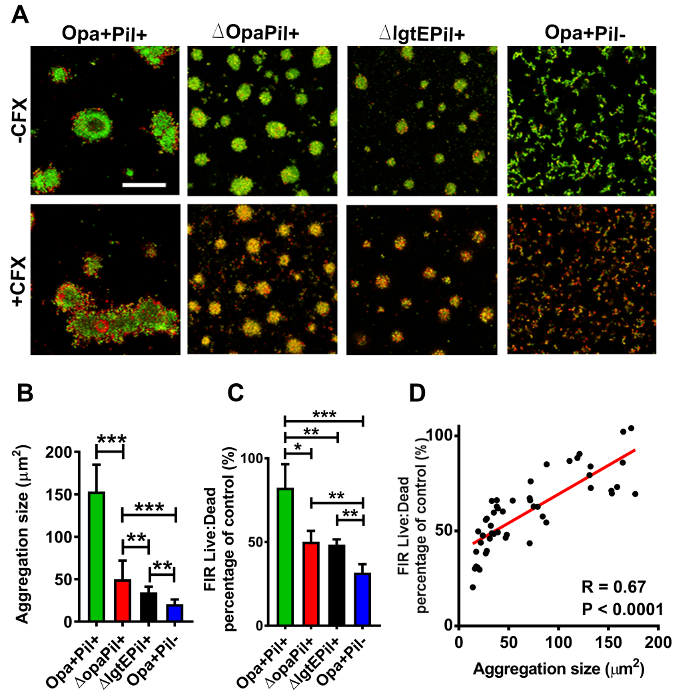
Figure 2: Representative data of live/dead bacteria distribution within aggregates under ceftriaxone treatment. (A) Pre- aggregated MS11Opa+Pil+, MS11ΔOpaPil+, MS11Opa+Pil-, or MS11ΔLgtEPil+ was either incubated in the presence or absence of 1 µg/mL ceftriaxone for 2 h. Aggregates were then stained to visualize viable (green) and dead (red) GC and visualized with confocal fluorescence microscope. Scale bar: 50 µm. Images were then analyzed for (B) aggregation size and (C) ratio of live-to-dead GC and (D) a correlation graph was then created. Shown are the average values (SD) obtained from > 40 images of three independent experiments. ***p < 0.001; **p < 0.01; *p < 0.05. This figure was previously published 24and is used with permission. Please click here to view a larger version of this figure.
