Noen reaktive oksygen arter (ROS) er i stand til å oksidere karbon doble bånd av DNA baser og noen karbonatomer i deoxyribose moiety, genererer oksidert baser og DNA strand pauser1. Som et negativt ladet molekyl rik på nitrogen og oksygen atomer, er DNA også et mål for Elektrofil grupper som covalently reagerer med nukleofil nettsteder (nitrogen og oksygen), noe som gir produkter som kalles DNA addukter2. Så, DNA addukter og oksidert DNA baser er eksempler på DNA lesjoner som er nyttige biomarkører for toksisitet vurdering av stoffer som er Elektrofil, generere reaktive elektrofiler ved biotransformasjon, eller indusere oksidativt stress1, 2i det. Selv om de modifiserte DNA baser kan fjernes fra DNA ved basen eller nukleotid forbrukeravgift reparasjon (BER eller NER), induksjon av en ubalanse mellom generering og fjerning av DNA lesjoner i favør av den tidligere fører til en netto økning på sine nivåer i DNA over tid3 . Resultatene er økningen av DNA mutasjon priser, redusert genuttrykk, og redusert protein aktivitet2,4, 5,6,7, effekter som er nært knyttet til utvikling av sykdommer. DNA mutasjoner kan påvirke ulike cellulære funksjoner, slik som celle signalering, celle syklus, Genova integritet, telomere stabilitet, epigenome, kromatin struktur, RNA skjøting, protein homeostase, metabolisme, apoptose, og celle differensiering8 ,9. Strategier for å forsinke celle mutasjon priser og kronisk sykdom utvikling (f. eks, kreft, nevrodegenerative sykdommer) passere gjennom kunnskap om mutasjon kilder, blant dem, DNA lesjoner og deres årsaker.
ROS generert endogenously i overkant, på grunn av forurensende eksponering, vedvarende betennelse, sykdom patofysiologi (f. eks, diabetes), etc., er viktige årsaker til biomolekyler Kader, inkludert DNA og lipid skade1. Som et eksempel, den svært reaktive hydroksyl radikale (OH) dannet fra H2O2 reduksjon av overgangen metall ioner (Fe2 +, Cu+) oksiderer DNA baser, DNA sukker moiety og flerumettede fettsyrer ved diffusjon-kontrollerte priser10. Blant 80 allerede karakterisert oksidert nucleobases3, den mest studerte en er 8-Oxo-7, 8-dihydroguanine (8-oxoGua) eller 8-Oxo-7, 8-dihydro-2′-deoxyguanosine (8-OxodGuo, figur 1), en lesjon som er i stand til å indusere gt transversions i pattedyrceller10,11. Det er dannet av mono elektronisk oksidasjon av Guanin, eller ved hydroksyl radikale eller singlet oksygen angrep av Guanin i DNA1. Flerumettede fettsyrer er andre viktige mål for svært reaktive oksidanter, for eksempel •Oh, som starter prosessen med lipid peroxidation1,12. Det gir opphav til fatty acid hydroperoxides som kan brytes ned til Elektrofil aldehyder og epoxyaldehydes, som malondialdehyde, 4-AHA-2-nonenal, 2, 4-decadienal, 4, 5-epoxy-(2E)-decenal, hexenal, acrolein, crotonaldehyde, som er stand til å danne mutagent exocyclic DNA addukter, slik som malondialdehyde-, propano-, eller etheno addukter1,12,13. Den etheno addukter 1,n2-etheno-2′-Deoxyguanosine (1,n2-ΕdGuo, figur 1) og 1,n6-etheno-2′-deoksyadenosinregionen (1,n6-εdAdo, figur 1 ) har blitt foreslått som potensielle biomarkører i patofysiologi av betennelse14,15.
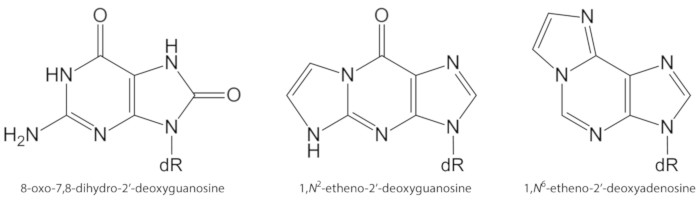
Figur 1. Kjemiske strukturer av DNA lesjoner kvantifisert i denne studien. dR = 2 ́-deoxyribose. Dette tallet er modifisert fra Oliveira et al.34. Vennligst klikk her for å se en større versjon av dette tallet.
Studier utført tidlig på 1980-tallet tillot sensitiv påvisning av 8-oxodGuo av høy ytelse flytende kromatografi koplet til elektrokjemiske deteksjon (HPLC-ECD). Kvantifisering av 8-oxodGuo av HPLC-ECD i flere biologiske systemer utsatt for oksiderende forhold førte til anerkjennelse av 8-oxodGuo som en biomarkør av oxidatively indusert base skade i DNA1,16. Selv om den er robust og tillater kvantifisering av 8-oxodGuo i det lave fmol området17, er HPLC-ECD-målinger avhengig av nøyaktigheten av analytt oppbevaringstid for analytt identifisering og kromatografi oppløsning for å unngå forstyrrelser i andre utvalgs bestanddeler. Som elektrokjemiske deteksjon krever bruk av salt (f. eks, kalium fosfat, natrium acetate) i den mobile fasen, vedlikehold av tilstrekkelig analytiske forhold behov rutine kolonne og utstyr rengjøring tid.
Alternativt, bruk av bakteriell DNA reparasjon enzym formamidopyrimidine DNA glycosylase (FPG) og, etterpå, menneskelige 8-oxoguanine glycosylase 1 (hOGG1), for påvisning og fjerning av 8-oxoGua fra DNA, dukket opp som en måte for induksjon av DNA alkali labilt Nettsteder. Den alkali labilt nettsteder er konvertert til DNA strand pauser og la svært høysensitiv indirekte kvantifisering av 8-oxoGua av alkalisk enkelt celle gel elektroforese (“komet analysen”). Den høye følsomheten og gjennomføringen av analysene uten behov for mobilnettet DNA utvinning er de viktigste fordelene med denne typen analysen. Det gir lavest steady-state nivåer av 8-oxoGua i DNA, typisk 7-10 ganger lavere enn nivåene innhentet av bioanalytical metoder basert på HPLC. Men det er en indirekte måling av 8-oxoGua og noen ulemper er mangelen på spesifisitet eller ukjent effektiviteten av reparasjon enzymer som brukes1,16,18.
Immunanalyser er andre sett av metoder som brukes for påvisning av 8-oxoGua1 og exocyclic DNA addukter, for eksempel 1,N6-dAdo og 1,n2-dGuo12. Til tross for følsomheten, en brist av bruk av antistoffer for påvisning av DNA lesjoner er mangelen på spesifisitet på grunn av kryss-reaktivitet til andre komponenter av biologiske prøver, inkludert normal DNA baser1,12. Exocyclic DNA addukter, inkludert 1,n6-DAdo og 1,n2-dGuo, kan også oppdages og kvantifisert av svært følsomme 32P-postlabeling analyser12. Den høye følsomheten til 32P-postlabeling tillater bruk av svært små mengder DNA (f. eks 10 μg) for påvisning av ca 1 epoxyaddukt per 1010 normal baser19. Men bruken av radio-kjemikalier, mangel på kjemisk spesifisitet og lav nøyaktighet er noen ulemper19,20.
En felles begrensning av metodene sitert ovenfor er den lave selektivitet eller spesifisitet for påvisning av de ønskede molekyler. I dette scenariet, HPLC koblet til electrospray ionisering tandem masse massespektrometri (HPLC-ESI-MS/MS og HPLC-MS3) utviklet seg som gullstandarden for kvantifisering av modifisert nucleosides i biologiske matriser, som DNA, urin, plasma og spytt 1 den andre , 19 andre priser , 20. fordeler med HPLC-ESI-MS/MS metoder er følsomheten (vanligvis i lav fmol rekkevidde) og den høye spesifisitet levert av i) den kromatografiske separasjon, II) den karakteristiske og kjente mønster av molekyl fragmentering inne i massen spektrometer kollisjons kammer, og III) nøyaktig måling av den valgte massen til lade ratio (m/z) i flere reaksjons overvåkingsmodus1,19. Bruken av isotopically merket interne standarder legger til fordel for rettelser for molekylet tap under DNA hydrolyse og analytt berikelse skritt, samt for forskjeller i analytt ionisering mellom prøvene. Den likeledes hjelpemidler inne identifikasjonen av det korrekt kromatografiske fjellpigg når mer enn ettall fjellpigg er gave1,12,19,20.
Adskillige metoder basert på HPLC-ESI-MULTIPLE SCLEROSIS/MULTIPLE SCLEROSIS ha blitt anvendt for kvantifisering av 8-oxodGuo, 1,N6-dAdo og 1,N2-dGuo inne DNA utdraget fra annerledes Biological prøvene12,15,20 ,21,22,23,24,25,26,27,28,29 . Fine partikler (PM2,5) bærer organiske og uorganiske kjemikalier som Polysykliske aromatiske hydrokarboner (PAH), Nitro-PAH, aldehyder, ketoner, kar bok syls syrer, quinolines, metaller og vannløselige ioner, som kan indusere betennelser og oksidativt stress, forhold som favoriserer forekomsten av biomolekyler Kader og sykdom30,31,32,33. Vi presenterer her validert HPLC-ESI-MS/MS metoder som ble med hell søkt om kvantifisering av 8-oxodGuo, 1,N6-DAdo og 1,N2-dGuo i lunge, lever og nyre DNA av A/J mus for vurderingen av effekter av ambient PM2,5 eksponering34.
