Certaines espèces d’oxygène réactif (ROS) sont capables d’oxyder des liaisons doubles de carbone de bases d’ADN et de certains carbones dans la portion désoxyribose, générant des bases oxydées et des ruptures de brin d’ADN1. En tant que molécule chargée négativement riche en atomes d’azote et d’oxygène, l’ADN est aussi une cible pour les groupes électrophiles qui réagissent de façon covalente avec les sites nucléophiles (azote et oxygène), donnant des produits qui sont appelés adduits d’ADN2. Ainsi, les adduits d’ADN et les bases d’ADN oxydées sont des exemples de lésions de l’ADN qui sont des biomarqueurs utiles pour l’évaluation de la toxicité des substances qui sont électrophiliques, génèrent des électrophiles réactifs lors de la biotransformation, ou induisent un stress oxydatif1, 2. les deux. Bien que les bases modifiées de l’ADN puissent être retirées de l’ADN par réparation d’excision de base ou de nucléotide (BER ou NER), l’induction d’un déséquilibre entre la génération et l’élimination des lésions de l’ADN en faveur de l’ancien conduit à une augmentation nette de leurs niveaux dans les heures supplémentaires de l’ADN3 . Les résultats sont l’augmentation des taux de mutation de l’ADN, l’expression génique réduite et l’activité protéique diminuée2,4,5,6,7, effets qui sont étroitement liés à la développement des maladies. Les mutations de l’ADN peuvent affecter diverses fonctions cellulaires, telles que la signalisation cellulaire, le cycle cellulaire, l’intégrité du génome, la stabilité du télomère, l’épigénome, la structure de la chromatine, l’épissage d’ARN, l’homéostasie protéique, le métabolisme, l’apoptose et la différenciation cellulaire8 ,9. Les stratégies visant à ralentir les taux de mutation cellulaire et le développement des maladies chroniques (p. ex. cancer, maladies neurodégénératives) traversent la connaissance des sources de mutation, parmi lesquelles les lésions de l’ADN et leurs causes.
Les ROS générés de façon endogène en excès, en raison de l’exposition aux polluants, de l’inflammation persistante, de la pathophysiologie des maladies (p. ex. le diabète), etc., sont des causes importantes de dommages causés par la biomolécule, y compris l’ADN et les lésions lipidiques1. À titre d’exemple, le radical hydroxyle hautement réactif (OH) formé à partir de H2O2 réduction par des ions métalliques de transition (Fe2 +, Cu+) oxyde les bases de l’ADN, la portion de sucre d’ADN et les acides gras polyinsaturés à la diffusion contrôlée taux de change10. Parmi les 80 déjà caractérisés nucléobases oxydées3, la plus étudiée est la 8-oxo-7, 8-dihydroguanine (8-oxoGua) ou 8-oxo-7, 8-dihydro-2 ‘-désoxyguanosine (8-oxodGuo, figure 1), une lésion qui est capable d’induire gt transversions dans cellules de mammifères10,11. Il est formé par l’oxydation mono-électronique de la guanine, ou par le radical hydroxyle ou l’attaque de l’oxygène sinéne de la guanine dans l’ADN1. Les acides gras polyinsaturés sont d’autres cibles importantes d’oxydants hautement réactifs, tels que •Oh, qui initie le processus de peroxydation lipidique1,12. Il donne lieu à des hydroperoxydes d’acides gras qui peuvent se décomposer en aldéhydes électrophiliques et en époxyaldéhydes, comme le malondialdéhyde, le 4-hydroxy-2-nonenal, le 2,4-décadienal, le 4,5-époxy-(2E)-décénal, l’Hexénal, l’acroléine, le crotonaldéhyde, qui sont capables de former des adduits d’ADN exocyclique mutagènes, tels que les adduits de malondialdéhyde-, Propano-ou étheno1,12,13. Les adduits d’étheno 1,n2-étheno-2′-désoxyguanosine (1,n2-εdguo, figure 1) et 1,n6-étheno-2 ‘-désoxyadénosine (1,n6-εdado, figure 1 ) ont été suggérés comme biomarqueurs potentiels dans la pathophysiologie de l’inflammation14,15.
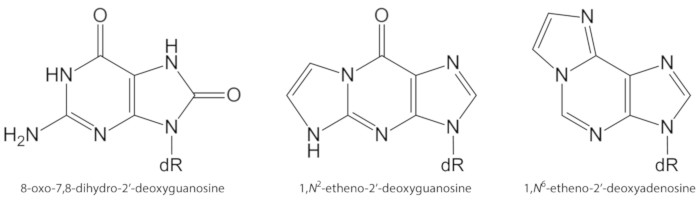
La figure 1. Les structures chimiques des lésions de l’ADN quantifiées dans la présente étude. dR = 2 ́-désoxyribose. Ce chiffre a été modifié à partir de Oliveira et al.34. S’il vous plaît cliquez ici pour voir une version plus grande de ce chiffre.
Des études réalisées au début des années 1980 ont permis la détection sensible du 8-oxodGuo par chromatographie liquide à haute performance couplée à la détection électrochimique (HPLC-ECD). La quantification du 8-oxodguo par HPLC-ECD dans plusieurs systèmes biologiques soumis à des conditions oxydantes a conduit à la reconnaissance du 8-oxodguo comme biomarqueur des lésions de base induites oxydativement dans l’ADN1,16. Bien que robustes et permettant la quantification du 8-oxodGuo dans la gamme faible de fmol17, les mesures HPLC-ECD reposent sur la précision du temps de rétention de l’analyte pour l’identification de l’analyte et sur la résolution de la chromatographie afin d’éviter les interférences de autres constituants de l’échantillon. Comme la détection électrochimique nécessite l’utilisation de sel (p. ex., phosphate de potassium, acétate de sodium) dans la phase mobile, le maintien de conditions analytiques adéquates nécessite un temps de nettoyage de la colonne et du matériel de routine.
Alternativement, l’utilisation de l’enzyme de réparation de l’ADN bactérien formamidopyrimidine DNA glycosylase (FPG) et, par la suite, de l’humain 8-oxoguanine glycosylase 1 (hOGG1), pour la détection et l’enlèvement de 8-oxoGua de l’ADN, est apparu comme un moyen pour l’induction de l’ADN labiles Sites. Les sites labiles alcalins sont convertis en ruptures de brin d’ADN et permettent la quantification indirecte très élevée sensible de la 8-oxoGua par électrophorèse alcaline de gel de cellule simple («dosage de comète»). La haute sensibilité et l’accomplissement des analyses sans besoin d’extraction d’ADN cellulaire sont les principaux avantages de ce type de dosage. Il donne les plus faibles niveaux stables de 8-oxoGua dans l’ADN, typiquement 7-10 fois plus bas que les niveaux obtenus par des méthodes bioanalytiques basées sur HPLC. Cependant, il s’agit d’une mesure indirecte de la 8-oxogua et certains inconvénients sont le manque de spécificité ou l’efficacité inconnue des enzymes de réparation utilisés1,16,18.
Les immunodosages sont d’autres méthodes utilisées pour la détection des adduits de l’ADN 8-oxoGua1 et exocyclique, tels que 1,n6-Dado et 1,n2-dguo12. Malgré la sensibilité, une lacune de l’utilisation des anticorps pour la détection des lésions de l’ADN est le manque de spécificité en raison de la réactivité croisée à d’autres composants d’échantillons biologiques, y compris les bases normales d’ADN1,12. Les adduits d’ADN exocycliques, y compris 1,n6-Dado et 1,n2-dguo, peuvent également être détectés et quantifiés par des essais de P-post-étiquetage 32hautement sensibles12. La sensibilité élevée de 32P-le post-étiquetage permet l’utilisation de très petites quantités d’ADN (par exemple, 10 μg) pour la détection d’environ 1 adduit par 1010 bases normales19. Cependant, l’utilisation de la radio-chimie, le manque de spécificité chimique et la faible précision sont quelques inconvénients19,20.
Une limitation partagée des méthodes citées ci-dessus est la faible sélectivité ou spécificité pour la détection des molécules souhaitées. Dans ce scénario, la HPLC couplée à la spectrométrie de masse en tandem par ionisation par ÉLECTROPULVÉRISATION (HPLC-ESI-MS/MS et HPLC-MS3) a évolué comme étalon-or pour la quantification des nucléosides modifiés dans les matrices biologiques, comme l’ADN, l’urine, le plasma et la salive le premier , le 19 , 20. les avantages des méthodes HPLC-ESI-MS/MS sont la sensibilité (typiquement dans la plage de fmol faible) et la haute spécificité fournie par i) la séparation chromatographique, II) le schéma caractéristique et connu de la fragmentation des molécules à l’intérieur de la masse chambre de collision du spectromètre, et III) la mesure exacte du rapport masse/charge sélectionné (m/z) en mode de surveillance de réaction multiple1,19. L’utilisation de normes internes étiquetées isotopiquement ajoute l’avantage de corrections pour les pertes de molécules au cours de l’hydrolyse de l’ADN et des étapes d’enrichissement de l’analyte, ainsi que pour les différences de l’ionisation de l’analyte entre les échantillons. Il aide également à identifier le pic chromatographique correct lorsque plus d’un pic est présent1,12,19,20.
Plusieurs méthodes basées sur HPLC-ESI-MS/MS ont été utilisées pour la quantification de 8-oxodguo, 1, n6-Dado et 1,n2-dguo dans l’ADN extrait de différents échantillons biologiques12,15,20 ,21,22,23,24,25,26,27,28,29 . Particules fines (PM2,5) portent des produits chimiques organiques et inorganiques, tels que les hydrocarbures aromatiques polycycliques (HAP), les nitro-HAP, les aldéhydes, les cétones, les acides carboxyliques, les quinolines, les métaux et les ions solubles dans l’eau, qui peuvent induire une inflammation et stress oxydatif, conditions qui favorisent l’apparition de dommages de biomolécule et de la maladie30,31,32,33. Nous présentons ici les méthodes validées HPLC-ESI-MS/MS qui ont été appliquées avec succès pour la quantification de 8-oxodGuo, 1,n6-Dado et 1,n2-dguo dans l’ADN pulmonaire, hépatique et rénale de souris A/J pour l’évaluation de la effets de l’exposition ambiante2,5 de PM34.
