Einige reaktive Sauerstoffarten (ROS) sind in der Lage, Kohlenstoff-Doppelbindungen von DNA-Basen zu oxidieren und einige Kohlenstoff in der Deoxyribose-Feuchtigkeit, wodurch oxidierte Basen und DNA-Strangbrüche 1 erzeugt werden. Als negativ geladenes, stickstoff-und Sauerstoffatome geladenes Molekül ist die DNA auch ein Ziel für elektrophile Gruppen, die mit den nukleophilen Standorten (Stickstoff und Sauerstoff) kovalent reagieren und Produkte liefern,die als DNA-Adduktoren bezeichnet werden. DNA-Addukte und oxidierte DNA-Basen sind also Beispiele für DNA-Läsionen, die nützliche Biomarker für die Toxizitätsbewertung von elektrophilen Substanzen sind, reaktive Elektrophilen bei Bioveränderung erzeugen oder oxidativen Stress erzeugen1, 2. Runde Obwohl die modifizierten DNA-Basen durch Basis-oder Nukleotid-Exzisionsreparatur (BER oder NER) aus der DNA entfernt werden können, führt die Induktion eines Ungleichgewichts zwischen der Erzeugung und Entfernung von DNA-Läsionen zu Gunsten der ersteren zu einer Nettoerhöhung ihrer DNA-Überstunden 3. Ergebnisse sind die Erhöhung der DNA-Mutationsraten, eine verminderte Genexpression und eineverminderte Proteinaktivität 2, 4,5, 6, 7,Effekte, die engmitder Entwicklung von Krankheiten. DNA-Mutationen können sich auf verschiedene zelluläre Funktionen auswirken, wie z. B. Zellsignalisierung, Zellzyklus, Genomintegrität, Telomerstabilität, Epigenom, Chromatin-Struktur, RNA-Splicing, Protein-Haffeestose, Stoffwechsel, Apoptose und Zelldifferenzierung8 ,9. Strategien zur Verlangsamung von Zellmutationsraten und chronischer Krankheitsentwicklung (z.B. Krebs, neurodegenerative Erkrankungen) durchlaufen das Wissen um die Mutationsquellen, darunter DNA-Läsionen und deren Ursachen.
ROS, die endogen im Übermaß erzeugt werden, aufgrund von Schadstoffexposition, hartnäckiger Entzündung, Krankheitspophysiologie (z.B. Diabetes), etc., sind wichtige Ursachen für Biomolekule-Schäden, einschließlichDNA und Fettschäden 1. Zum Beispiel oxidiert das hochreaktive Hydroxylradikal (OH), das sich aus der H2O 2-Reduktion durch Übergangs-Metallionen (Fe2 +, Cu+) gebildet hat, die DNA-Basen, die DNA-Zuckergerüchte und die mehrfach ungesättigten Fettsäuren bei diffusionsgesteuerten Fettsäuren. Sätze10. Unter den 80 bereits charakterisierten oxidierten Nukleobasen3, Die am meisten untersuchte ist 8-oxo-7,8-Dihydroguanin (8-oxoGua) oder 8-oxo-7,8-Dihydro-2 ‘-Deoxyguanosin (8-oxodGuo , Abbildung1), eine Läsion, die in der Lage ist, GT-Transformatoren in induzieren Säugetierzellen10,11. Es wird durch die monoelektronische Oxidation von Guanin oder durch Hydroxylradikal oder Singlet-Sauerstoff-Attacke von Guanin in DNA1 gebildet. Polyungesättigte Fettsäuren sind weitere wichtige Ziele hochreaktiver Oxidantien, wie zum Beispiel • OH, die den Prozess der Fettperoxidation1,12diGang setzen. Es entstehen Fettsäure-Hydroperoxide, die zu elektrophilen Aldehyden und Epoxyaldehyden zersetzen können, Wie Malondialdehyd, 4-hydroxy-2-nonenal, 2,4-deadienal, 4,5 epoxy-(2E)-dezental, hexenal, Akrolein, Krotonaldehyd, die In der Lage, mutagene exozyklische DNA-Addukte wie Malondialdehyde-, Propano-oder Etheno-Addukte 1,12,13zu bilden. Das Etheno Addukte 1,N2-etheno-2 ‘-deoxyguanosine (1,N2-DGuo , Abbildung 1) und 1,N6-etheno-2 ‘-deoxyadenosine (1,N6-madAdo, Abbildung 1 ) wurden als potenzielle Biomarker in der Pathophysiologie der Entzündung14,15vorgeschlagen.
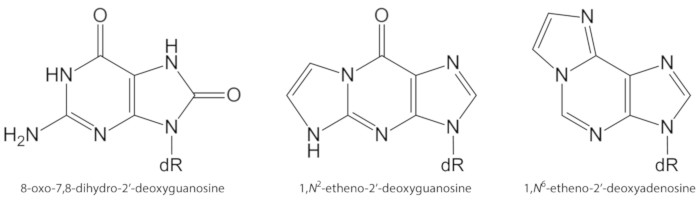
Bild 1. Chemische Strukturen der DNA-Läsionen, die in der vorliegenden Studie quantifiziert wurden. DR = 2 ́-deoxyribose. Diese Figur wurde von Oliveira et al. 34 geändert. Bitte klicken Sie hier, um eine größere Version dieser Figur zu sehen.
Studien, die in den frühen 1980er Jahren durchgeführt wurden, ermöglichten die sensible Erkennung von 8-oxodGuo durch eine hochleistungsfähige Flüssigchromatographie in Verbindung mit elektrochemischer Erkennung (HPLC-ECD). Die Quantifizierung von 8-oxodGuo durch die HPLC-ECD in mehreren biologischen Systemen, die oxidierenden Bedingungen unterworfen waren, führte zur Erkennung von 8-oxodGuo als Biomarker oxidativ induzierter Grundschäden in DNA1,16. Obwohl robust und die Quantifizierung von 8-oxodGuo im niedrigen Fmol-Bereich 17 ermöglicht, setzen die HPLC-EKD-Messungen auf die Genauigkeit der Analyte-Retendesszeit zur Analyteerkennung und auf die Chromatographie-Auflösung, um Störungen von Andere Stichproben. Da die elektrochemische Erkennung den Einsatz von Salz (z.B. Kaliumphosphat, Natriumacetat) in der mobilen Phase erfordert, benötigt die Aufrechterhaltung adäquater Analysebedingungen eine routinemäßige Säulen-und Gerätereinigungszeit.
Alternativ ist die Verwendung des bakteriellen DNA-Reparaturenzyms formamidopyrimidine DNA Glycosylase (FPG) und, Danach entstand menschliches 8-oxoguanin-Glykosylase 1 (hOGG1) zur Erkennung und Entfernung von 8-oxoGua aus der DNA als Mittel zur Induktion von DNA-Alkali labile Websites. Die Alkali-Labillierungsstellen werden in DNA-Strang-Breaks umgewandelt und ermöglichen die sehr hohe empfindliche indirekte Quantifizierung von 8-oxoGua durch alkalische Einzelzellen-Gel-Elektrophorese (“Kometenuntersuchen”). Die hohe Empfindlichkeit und die Durchführung der Analysen ohne zelluläre DNA-Extraktion sind die Hauptvorteile dieser Art von Test. Es gibt den niedrigsten Stand-Zustand-Wert von 8-oxoGua in der DNA, in der Regel 7-10 Mal niedriger als die, die durch bioanalytische Methoden auf der Basis von HPLC erhalten. Es handelt sich jedoch um eine indirekte Messung von 8-oxoGua und einige Nachteile sind die fehlende Spezifität oder die unbekannte Effizienz der verwendeten Reparaturenzyme1,16,18.
Immunoassays sind weitere Methoden, die zur Erkennung von 8-oxoGua1 und exozyklischen DNA-Addukten verwendet werden, wie1, N6-dAdo und 1,N2-dGuo12. Trotz der Empfindlichkeit ist ein Mangel an Antikörpern zur Erkennung von DNA-Läsionen der Mangel an Spezifität aufgrund von Kreuzreaktionen auf andere Bestandteile biologischer Proben, einschließlich der normalenDNA-Basen 1,12. Die exozyklischen DNA-Addukte, darunter 1,N6-dAdo und1, N2-dGuo, können auch durch hochsensible 32P-Postbeschriftungsuntersuchungen 12 erkanntund quantifiziert werden. Die hohe Empfindlichkeit von 32P-Postbeschriftung ermöglicht die Verwendung von sehr geringen Mengen an DNA(z.B. 10 μg) zur Erkennung von etwa 1 Addukt pro 1010 normalen Basen 19. Allerdings sind der Einsatz von Radio-Chemikalien, mangelnde chemische Spezifität und geringe Genauigkeit einige Nachteile 19,20.
Eine gemeinsame Einschränkung der oben genannten Methoden ist die geringe Selektivität oder Spezifität für die Erkennung der gewünschten Moleküle. In diesem Szenario entwickelte sich HPLC in Verbindung mit der Elektrospray-Ionisierung der Massenspektrometrie (HPLC-ESI-MS/MS und HPLC-MS3) zu dem Goldstandard für die Quantifizierung von modifizierten Nukleoside in biologischen Matrizen, wie DNA, Urin, Plasma und Speichel. 1 , 19 , 20. Vorteile der HPLC-ESI-MS/MS-Methoden sind die Empfindlichkeit (typischerweise im niedrigen Fmol-Bereich) und die hohe Spezifität, die i zur Verfügung stellt, die chromatographische Trennung, ii) das charakteristische und bekannte Muster der Molekül-Fragmentierung innerhalb der Masse Spektrometer-Kollisionskammer, und iii) die genaue Messung des ausgewählten Massen-bis Ladungsverhältnisses (m/z) im Multitreaktionsüberwachungsmodus 1,19. Der Einsatz isotopisch beschrifteter interner Standards erhöht den Vorteil von Korrekturen für Molekülverluste während der DNA-Hydrolyse-und Analytenanreicherungsschritte sowie für Unterschiede bei der Analytitisierung zwischen Proben. Es hilft auch bei der Identifizierung der richtigen chromatographischen Spitze, wenn mehr als ein Peak ist1,12,19,20.
Für die Quantifizierung von 8-oxodGuo,1,N6-dAdo und 1,N 2-dGuo in DNA, die aus verschiedenen biologischen Proben gewonnen wurde, wurden mehrere Methoden verwendet, die auf HPLC-ESI-MS/MSbasieren. ,21,22, 23,24, 25, 26, 27,28,29 . Feinstaub (PM 2.5) trägt organische und anorganische Chemikalien, wie polyzyklische aromatische Kohlenwasserstoffe (PAK), Nitro-PAHs, Aldehyde,Ketone, Carboxylsäuren, Quinoline, Metalle und wasserlösliche Ionen, die Entzündungen verursachen können und Oxidativer Stress, Bedingungen, die das Auftreten von Biomolekule-Schäden und Krankheit30,31,32,33begünstigen. Wir stellen hier validierte HPLC-ESI-MS/MS-Methoden vor, die erfolgreich für die Quantifizierung von 8-oxodGuo,1, N6-dAdo und 1,N2-dGuo in Lungen-, Leber-und Nieren-DNA von A/J-Mäusen zur Beurteilung der Die Auswirkungen der UmgebungsbelastungPM2.5 34.
