Vesicle production
Figure 1 shows transmission electron microscopy (TEM) images of vesicles prepared with different swelling solutions and the glass beads method (also see Vogele et al.11). For the sample in Figure 1A, only PBS was used as swelling solution to prove the formation of vesicles and to determine their size. When TX-TL was used as swelling solution (Figure 1B), the vesicles also formed. Dynamic light scattering (DLS) measurements were performed to show that the glass beads method has an effect on vesicle formation. Figure 2 depicts the measured intensity autocorrelation curves of an EF sample prepared without glass beads in PBS (blue) and an EF sample prepared with the glass beads method with PBS as the swelling solution (red). The sizes were calculated from a cumulant fit25 and resulted in a diameter of 134 nm with a polydispersity of 25% when the vesicles were prepared without the glass beads method. When the glass beads were used, the cumulant fit resulted in a diameter of 168 nm with a polydispersity of 21%.
In vesiculo transcription11
Figure 3 shows the fluorescence intensity profile over time for the transcription of the dBroccoli aptamer inside the EF vesicles (green). As a negative control, DNase I was added to the swelling solution and thus also incorporated during vesicle formation (blue). The measurements were performed in fluorescence plate reader with excitation at 480 nm and emission at 520 nm.
In vesiculo protein expression11
Figure 4 shows the fluorescence intensity of two fluorescent proteins which were expressed inside the ELP vesicles using a fluorescence plate reader. Excitation was carried out 500 nm and emission at 520 nm. Transcription-translation of mVenus (Figure 4A) was performed to investigate expression dynamics and expression level of proteins in the ELP vesicle. It is important to note that after vesicle formation, the contents of the vesicles and outer solution are the same. Hence, to suppress protein expression outside of the vesicle, the antibiotic kanamycin was added to the exterior solution. As a control, kanamycin was also added to the swelling solution, in which case protein expression inside of the vesicle was suppressed. This further indicates that the small molecule kanamycin stays inside the vesicle and suggests that the membrane is not permeable for this molecule over the time scale of these experiments. If kanamycin diffused through the membrane, the mVenus expression would not have been suppressed and at 5 h, the fluorescence would be higher. In the second experiment, the expression of YPet (Figure 4B) was carried out. Both proteins were chosen because they exhibit a faster maturation time than, for instance, GFP. Furthermore, the T7 promoter was used for mVenus transcription and a constitutive promoter used for YPet transcription to show that both inducible and continuous expression are possible.
FRET assay11
Figure 5A shows the results of the FRET assay performed to demonstrate ELP incorporation into the membrane. Therefore, the vesicles were produced using a mixture of two equally concentrated fluorophore-peptide constructs. These were Cy3-EF (donor) and Cy5-EF (acceptor). At time t = 0, the starting FRET level was measured. The signals of the donor and the FRET signal depend on the mean distance between the dyes in the membrane. Upon expression of the membrane ELPs, additional peptides incorporate into the membrane, which increases the average distance between the FRET pairs. The latter was measured through the increase in donor fluorescence and a decrease of the FRET signal at time t = 2.5 h. Figure 5B shows the negative control. Here, kanamycin was added to the swelling solution before vesicle formation. Kanamycin suppresses protein expression; therefore, no change in FRET was visible. It is important to note that this assay only shows EF incorporation.
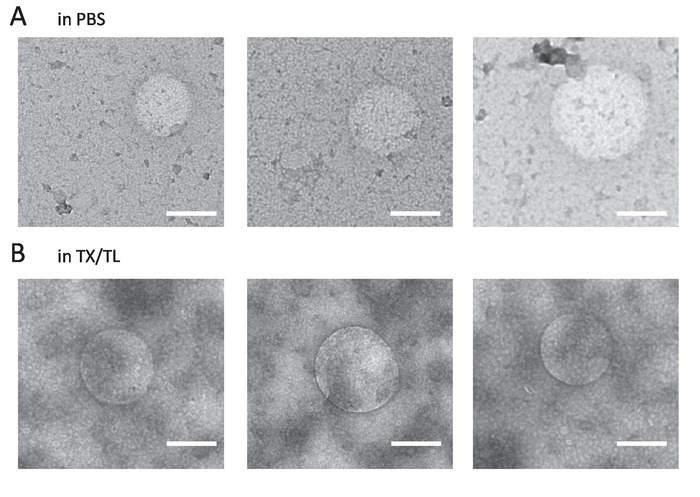
Figure 1: Representative TEM images. (A) EF vesicles formed in the swelling solution PBS. (B) EF vesicles formed in the swelling solution TX-TL. Scale bars = 200 nm. Please click here to view a larger version of this figure.
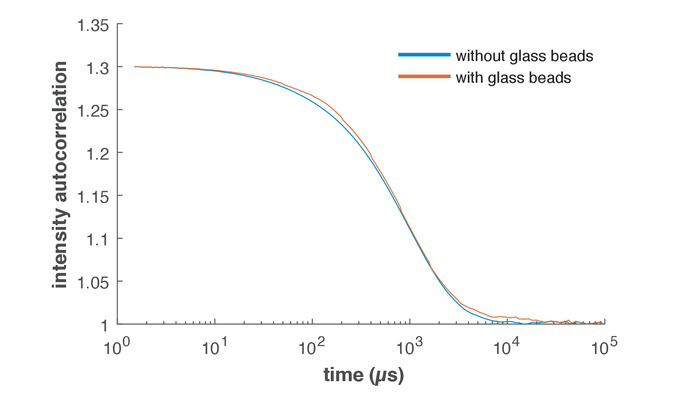
Figure 2: Representative DLS data. Intensity autocorrelation curves measured by DLS. The blue curve depicts EF peptides in PBS prepared without the glass beads method. The red curve depicts an EF solution produced with the glass beads method and PBS as the swelling solution. Please click here to view a larger version of this figure.
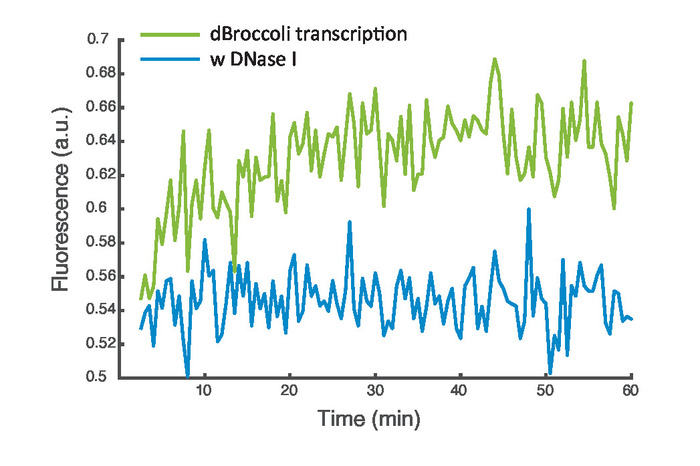
Figure 3: dBroccoli transcription. Representative data of the transcription of the dBroccoli aptamer inside EF vesicles. The green curve depicts the fluorescence signal and the blue curve the control measurement with encapsulated DNase I. Please click here to view a larger version of this figure.
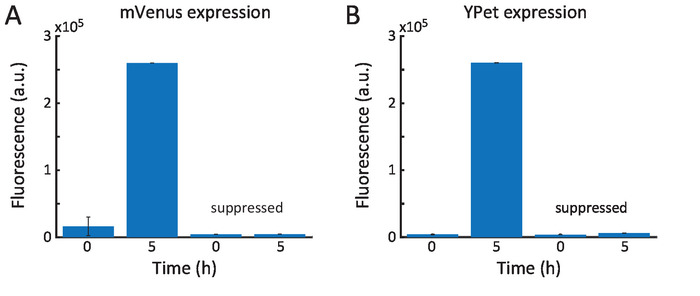
Figure 4: Protein expression inside peptide vesicles. (A) A plasmid encoding mVenus and the TX-TL expression mix were encapsulated in the ELP vesicles. After approximately 5 h, the fluorescence saturated. When the antibiotic kanamycin was encapsulated as well, no fluorescence was observed. (B) Similar results were obtained upon encapsulation of a YPet plasmid. Error bars represent the standard deviation of the measured values at the indicated time during a time frame of 15 min. Please click here to view a larger version of this figure.
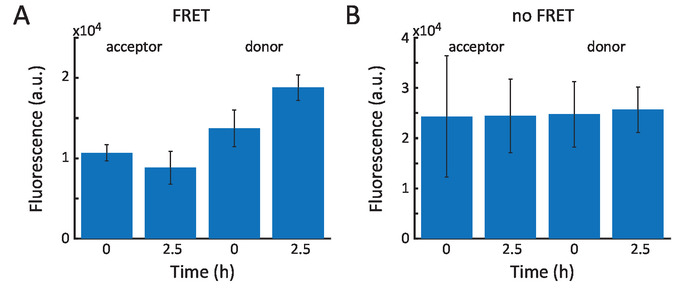
Figure 5: FRET assay. (A) Starting fluorescence levels at time t = 0 for donor emission and FRET signal (acceptor emission). At t = 2.5 h, the donor showed increased emission, and the FRET signal decreased. (B) When only hydrophilic ELP were expressed inside the vesicles, the FRET signal and donor emission stayed constant. Error bars represent the standard deviation of the measured values at the indicated time during a time frame of 15 min. Please click here to view a larger version of this figure.
Supplemental File: Contains all plasmid sequences resp. gene sequences. Please click here to download this file.
