


Summary
Vi præsenterer en metode til at fastsætte bestøvningskravene for abrikos(Prunus armenienca L.) sorter, der kombinerer bestemmelse af selv-(in)kompatibilitet ved fluorescensmikroskopi med identifikation af S-genotype ved PCR-analyse.
Abstract
Selv-uforenelighed i Rosaceae bestemmes af en Gametophytic Self-Incompatibility System (GSI), der hovedsagelig kontrolleres af den multiallelic locus S. I abrikos er det stadig vigtigere at bestemme selv- og inter-(in)kompatibilitetsforhold, da frigivelsen af et betydeligt antal nye sorter har resulteret i en stigning i sorter med ukendte bestøvningskrav. Her beskriver vi en metode, der kombinerer bestemmelse af selv-(in)kompatibilitet ved håndbestøvninger og mikroskopi med identifikation af S-genotype ved PCR-analyse. For selv-(in) kompatibilitet bestemmelse, blomster på ballon fase fra hver sort blev indsamlet i marken, hånd-bestøvet i laboratoriet, fast, og farves med anilin blå til observation af pollen rør adfærd under fluorescens mikroskopi. Med henblik på etablering af uforenelighedsrelationer mellem sorter blev DNA fra hver sort Sudvundet af unge blade, og S-alleler blev identificeret ved PCR. Denne fremgangsmåde gør det muligt at etablere uforenelighed grupper og belyse uforenelighed relationer mellem sorter, som giver en værdifuld information til at vælge egnede bestøvere i udformningen af nye plantager og til at vælge passende forældre i avlsprogrammer.
Introduction
Selv-uforenelighed er en strategi for blomstrende planter for at forhindre selvbestøvning og fremme outcrossing1. I Rosaceae bestemmes denne mekanisme af et Gametophytic Self-Incompatibility System (GSI), der hovedsageligt styres af den multiallelic locus S2. I den stil, RNase genet koder S-stylar determinant, en RNase3, mens en F-box protein, som bestemmer S-pollen determinant, er kodificeret af SFB genet4. Samspillet mellem sig selvi kompatibilitet finder sted gennem hæmning af pollenrørvækst i stil med at forhindre befrugtning af ovule5,6.
I abrikos, en sort fornyelse har fundet sted på verdensplan i de sidste to årtier7,8. Denne indførelse af et stort antal nye sorter, fra forskellige offentlige og private avlsprogrammer, har resulteret i stigningen af abrikos sorter med ukendte bestøvning krav8.
Forskellige metoder er blevet anvendt til at bestemme bestøvning krav i abrikos. På marken kan selv-in-kompatibilitet etableres ved kontrollerede bestøvninger i burtræer eller i kastrerede blomster og derefter registrere procentdelen af frugtsæt9,10,11og12. Desuden er kontrollerede bestøvninger blevet udført i laboratoriet ved semi-in vivo kultur af blomster og analyse af pollen rør adfærd under fluorescens mikroskopi8,13,14,15,16,17. For nylig, molekylære teknikker, såsom PCR analyse og sekventering, har gjort det muligt karakterisering af uforenelighed relationer baseret på studiet af RNase og SFB gener18,19. I abrikos er der rapporteret 33 S-alleler (S1 til S20, S22 til S30, S52, S53, Sv, Sx), herunder en allel relateret til selvkompatibilitet (Sc)12,18,20,21,22,23,24. Indtil nu er 26 inkompatibilitetsgrupper blevet ukket ned i denne art i henhold til S-genotype S8,9,,17,25,26,27. Kultiønder med Sde samme S-alleler er indbyrdes uforenelige, mens sorter med mindst én anden S-allelog derfor fordelt i forskellige uforenelige grupper er indbyrdes kompatible.
For at definere kravene til bestøvning af abrikoskultstoffer beskriver vi en metode, der kombinerer bestemmelse af selv-(in)kompatibilitet ved fluorescensmikroskopi med identifikation af S-genotype ved PCR-analyse i abrikoskulterer. Denne fremgangsmåde gør det muligt at etablere uforenelighedsgrupper og belyse uforenelighedsrelationer mellem sorter.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Selv-(in)kompatibilitet bestemmelse
- Prøve blomsterne i marken. Det er nødvendigt at indsamle blomsterne på ballonstadiet (Figur 1A), svarende til fase 58 på BBCH-skalaen for abrikos28, for at undgå uønsket tidligere bestøvning.
-
Selv- og krydsbestøvninger i laboratoriet
- Fjern anthers af blomsterne på ballonstadiet og læg dem på et stykke papir til at tørre ved laboratorietemperatur.
- Efter 24 timer sigtes pollenkernerne ved hjælp af en finmasket (0,26 mm) (figur 1B).
- Emasculate en gruppe på 30 blomster på samme ballon fase for hver selvbestøvning og krydsbestøvning og placere støvveje på blomsterskum i vand ved laboratorietemperatur (Figur 1C).
- Hånd bestøver støvveje ved hjælp af en pensel med pollen fra blomster af samme sort 24 timer efter emasculation. Desuden bestøver et andet sæt støvveje af hver sort med pollen fra blomster af en kompatibel bestøver som kontrol (Figur 1D).
- Efter 72 timer fastgøres støvvejene i en fiksativ opløsning af ethanol/eddikesyre (3:1) i mindst 24 timer ved 4 °C29. Kassér derefter fiksativt og tilsæt 75% ethanol, der sikrer, at prøverne er helt nedsænket i opløsningen. Prøverne kan opbevares i denne opløsning ved 4 °C indtil brug8,17,30,31,32.
-
Evaluering af pollen levedygtighed gennem in vitro pollen spiring
- Til fremstilling af spiremediet vægtes 25 g saccharose, 0,075 g borsyre (H3BO3) og 0,075 g calciumnitrat (Ca(NR3)2)33.
- Komponenterne i mediet tilsættes i 250 ml destilleret vand og opløses helt.
- Størkne mediet tilsætning 2 g agarose og blandes ved hvirvlende.
- Kontroller mediets pH-værdi ved hjælp af en pH-måler, og juster værdien til 7,0 med NaOH- eller HCl-opløsning.
- Autoklave blandingen til at sterilisere mediet.
- Efter autoklavering, køle ned mediet og distribuere det i petriskåle i en steril laminar flow hætte.
- Pollenkornene af de samme sorter, der anvendes til de kontrollerede bestøvninger i det størknede pollenspirationsmedium, spredes, og de observeres under mikroskopet efter 24 timer6.
BEMÆRK: For at sterilisere laminar-flowhætten skal overfladen rengøres med 70 % ethanol og tændes for UV-lampen i løbet af 10 min. - Petriskålene opbevares i køleskab ved 4 °C indtil brug.
-
Observationer af mikroskopi
- Pistils pistils tre gange for 1 time med destilleret vand og lad dem i 5% natriumsulfit ved 4 °C. Efter 24 timer, autoklave dem på 1 kg/cm2 i løbet af 10 min i natriumsulfit at blødgøre væv34.
- Placer autoklatrede støvveje over et glas dias og ved hjælp af en skalpel, fjerne trichomes omkring æggestokkene for at få en bedre visualisering af pollen rør. Derefter squash pistils med et dækglas.
- Der fremstilles 0,1% (v/v) anilinblå plet: Bland 0,1 ml anilinblå i 100 ml 0,1 N kaliumphosphat-tribasic (K3PO4). Påfør en dråbe anilin blå over forberedelserne til at plette callose depositions under pollen rør vækst.
- Overhold pollenrørene langs stilen ved et mikroskop med UV-epifluorescens ved hjælp af 340-380 bandpass og 425 longpass filtre.
2. DNA-ekstraktion
- Prøve 2-3 blade i feltet i foråret. Det anbefales at prøve bladene på unge stadier, da dna-fremstillet er af højere kvalitet og lavere niveauer af phenolforbindelser sammenlignet med gamle blade.
- Uddrag Genomisk DNA efter de trin, der er beskrevet i et kommercielt tilgængeligt kit (se Tabel over Materialer).
- Analysere mængden og kvaliteten af DNA-koncentrationer ved hjælp af UV-vis spektrofotometer (260 nm).
3. SS-allel identifikation
-
Opsætning af PCR-reaktioner
- Der fremstilles en 50 ng/μL fortynding i destilleret vand i hver DNA-ekstraktionsprøve.
- Tø pcr-reagenserne langsomt ud og opbevar dem på is. Lad DNA-polymerase i fryseren, indtil det er nødvendigt.
- Forbered forstærkningsreaktionerne ved hjælp af de forskellige kombinationer af primere. Opret PCR-reaktionssammensætningen ved at kombinere komponenterne i tabel 1. Vortex PCR reaktion mix godt og distribuere den mængde, der er angivet for de forskellige kombinationer af primere til hver brønd af PCR plade. Derefter tilsættes 1 μL af DNA-fortynding i hver brønd.
- Pcr-pladen anbringes i termocycleren, og det tilsvarende PCR-program, der er vist i tabel 1,køres .
- Analyser de forstærkede fragmenter. Der er hovedsagelig to forskellige måder at analysere PCR forstærkede fragmenter: kapillær elektroforese (CE) med fluorescerende-mærket primere eller som visualisere ampliconer af agarose gel elektroforese med ikke-mærkede primere.
-
Kapillær elektroforese
- For at fremstille belastningsbufferen blandes 35 μL deioniseret formamid med 0,45 μL mærket størrelsesstandard. Vortex reagenset til at blande godt, og derefter dispensere 35,5 μL i brønden af læserpladen.
- Der tilsættes 1 μL pcr-produktet i brønden. Hertil kommer, tilføje en dråbe mineralsk olie for at forhindre vand fordampning.
- Klargør separationspladen til at tilføje separationsbuffer.
- Brug den kommercielle software, der følger med genanalysatoren (se Tabel over Materialer). Opret en ny prøveplade, og gem prøvenavnene for alle brønde på pladen.
- Vælg analysemetoden. I dette tilfælde skal prøverne denatureres ved 90 °C i 120 s, injiceres ved 2,0 kV for 30 s og adskilles ved 6,0 kV i 35 min.
- Sæt de to plader ind i genanalysatoren. Fyld kapillær array med destilleret vand.
- Ilæg den patenterede lineære polyacrylamid (LPA) gel. Endelig skal du klikke på Kør.
-
Gel elektroforese
- Der fremstilles en 1% agarosegel tilsætning af 1,5 g molekylærbiologikvalitet agarose i 150 ml 1x TAE (Tris-aceta-EDTA) elektroforese, der kører buffer (40 mM Tris, 20 mM eddikesyre og 1 mM EDTA ved pH 8.0). Agarose opløses ved mikrobølgeopvarmning i 2-3 min.
- For at visualisere DNA, tilsættes 4 μL af en nukleinsyre plet (se Tabel af materialer) og blandes forsigtigt.
- Tilføj en gel kam, med tilstrækkelige brønde til stiger, kontrol og prøver, i en gel bakke. Hæld derefter langsomt blandingen i midten af gelbakken og undgå bobler.
- Lad gelen køle af i 30-45 minutter ved stuetemperatur, indtil gelen er helt størknet. Indfør gelen i elektroforesekammeret, fjern gelkammen og fyld kammeret med nok 1x TAE-buffer til at dække gelen.
BEMÆRK: Kontroller gelens placering. Brøndene skal placeres tæt på den negative pol, da negativt ladet DNA vandrer mod katoden. - Der tilsættes 5 μL læssebuffer (0,1 % (v/v) bromophenolblå) til PCR-produkterne, og der blandes godt.
- For at vurdere størrelsen af båndene, belastning 5 μL AF DNA molekylvægt stigen (se Tabel af Materialer).
- Læg prøverne i gelens ekstra brønde.
- Når alle prøverne og DNA-molekylvægtsstigen er lastet, skal gelen køres ved 90 V i 1-1,5 timer, indtil den blå farvestoflinje er ca. 75 % gelens længde.
- Visualiser båndene i en transilluminator for nukleinsyrer.
-
Kapillær elektroforese
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Bestøvningsundersøgelser i abrikos kræver brug af blomster i senballonstadiet en dag før antese (Figur 1A). Denne fase betragtes som den mest gunstige for både pollen og støvveje samling, da blomsterstrukturer er næsten modne, men anther dehiscence har endnu ikke fundet sted. Dette forhindrer interferens fra uønsket pollen, ikke kun af pollen fra samme blomst, men også fra andre blomster, da de lukkede kronblade hindrer ankomsten af insekter, der transporterer ekstern pollen. Pollenkornene sigtes let gennem et finmasket net (Figur 1B) fra dehiscerede anthers, der tidligere er anbragt på et stykke papir i 24 timer ved stuetemperatur eller med let ekstra varme. Ligeledes er støvveje fremstillet af blomster på ballonstadiet efter fjernelse af kronblade, bægerblade og støvdragere ved hjælp af pincet eller fingernegle (Figur 1C). Pistils kan være selvbestøvende og krydsbestøvet med en fin børste (Figur 1D).
De hermafroditiske blomster af abrikos har fem mørkerøde bægerblade, fem hvide kronblade (Figur 1A), et enkelt pistil (Figur 2A) og 25-30 støvdragere. Den pistil har tre hovedstrukturer: stigmatisering, stil og æggestok. Æggestokkene har to æggeser, og befrugtning af mindst én af dem er nødvendig for frugt indstilling. Under bestøvningen overfører insekter, hovedsagelig bier, pollenkorn til stigmatisering (figur 1A), hvor de spirer (figur 2B) inden for 24 timer efter bestøvning. En pollen rør er fremstillet af hver spirende pollen korn, som vokser gennem støvveje strukturer for at nå æggestokken efter 3-4 dage og befrugte en af de to æggehuse efter omkring 7 dage. I selv uforenelige sorter, hvor S-allelen af pollenkornet er den samme som en af de to af støvdyrene, stopper pollenrøret med at vokse i den øverste stil, hvilket forhindrer befrugtning ( figur2C). Pollenrørene fra en kompatibel sort med en anden S-allel kan dog vokse gennem stilen (Figur 2D), nå æggestokkene (figur 2 S E) og gøde en af de to æggefjer.
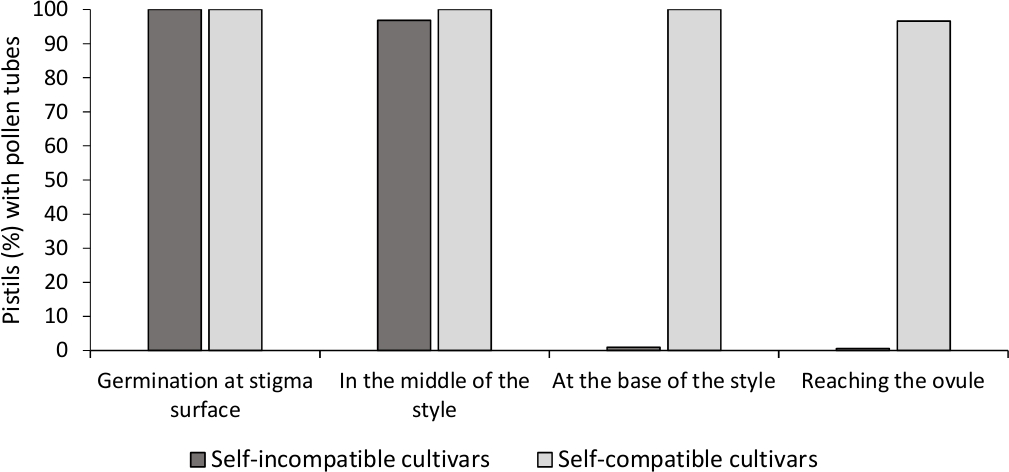
Analysen af in vitro pollenspiration viste god pollen levedygtighed i alle de sorter analyseret her, da de fleste pollen rør var længere end længden af pollen korn efter 24 timer i kulturmediet. Der blev observeret spirede pollenkerner ved stigmatiseringsoverfladen (figur 2B) i støvveje fra alle bestøvninger, hvilket indikerer tilstrækkelig bestøvning (figur 3).
For at bestemme selv-(in)kompatibilitet for hver sort, pollen rør adfærd i selv- og krydsbestøvninger udført i laboratorie-kontrollerede forhold blev observeret under fluorescens mikroskopi. Pollen rør vækst blev registreret langs den stil i alle de støvveje undersøgt. Kultièringer blev betragtet som selvkompatibile, da pollenrørsvækst blev arresteret i stilen i de fleste selvbestøvede støvveje (figur 2C, figur 3) og selvkompatible, når mindst et pollenrør nåede bunden af stilen i de fleste af de undersøgte støvveje (figur 2E, figur 3).
Undersøgelsen af S-locus ved PCR analyse tilladt karakterisering af S-genotype af hver sort. For det Sførste blev S-allelerne identificeret ved forstærkning af den første S-RNase intron ved hjælp af primerne SRc-F/SRc-R (tabel 2). Størrelsen af de forstærkede fragmenter blev analyseret ved kapillær elektroforese (Figur 4A) og blev brugt til at klassificere genotyper analyseret i deres tilsvarende uforenelighed gruppe (IG) (Tabel 3).
Nogle par alleler, såsom S1 og S7 eller S6 og S9, viste lignende fragment størrelser for den første intron. Differentieringen af disse alleler blev således foretaget ved at forstærke en region af RNase's anden intron med primerne Pru-C2/PruC4R, SHLM1/SHLM2 og SHLM3/SHLM4 (tabel 2). PruC2/PruC4R-primerkombinationen blev anvendt til at skelne mellem S6 og S9. For S6blev der forstærket et fragment på 1300 bp, mens der blev observeret et fragment på ca. 700 bp for S9-allelen ( figur4B, tabel 3). De specifikke primere SHLM1/SHLM2 og SHLM3/SHLM4 forstærkede et fragment på ca. 650 bp i S1 allel og 413 bp i S7 allel ( Figur4C, Tabel 3).
De primere AprFBC8-(F/R), der forstærker de variable V2- og HVb-områder i SFB-genet, blev anvendt til at skelne mellem Sc og S8-alleler, da begge alleler udviser identisk RNase-sekvens. S8-allelen viste et PCR-fragment på ca. 150 bp, mens et 500 bp-fragment svarede til Sc allelen (figur 4D, tabel 3). Da SS-genotypen blev fastlagt for alle sorter, blev selv uforenelige sorter tildelt deres tilsvarende uforenelighedsgrupper baseret på deres S-alleler( tabel3).
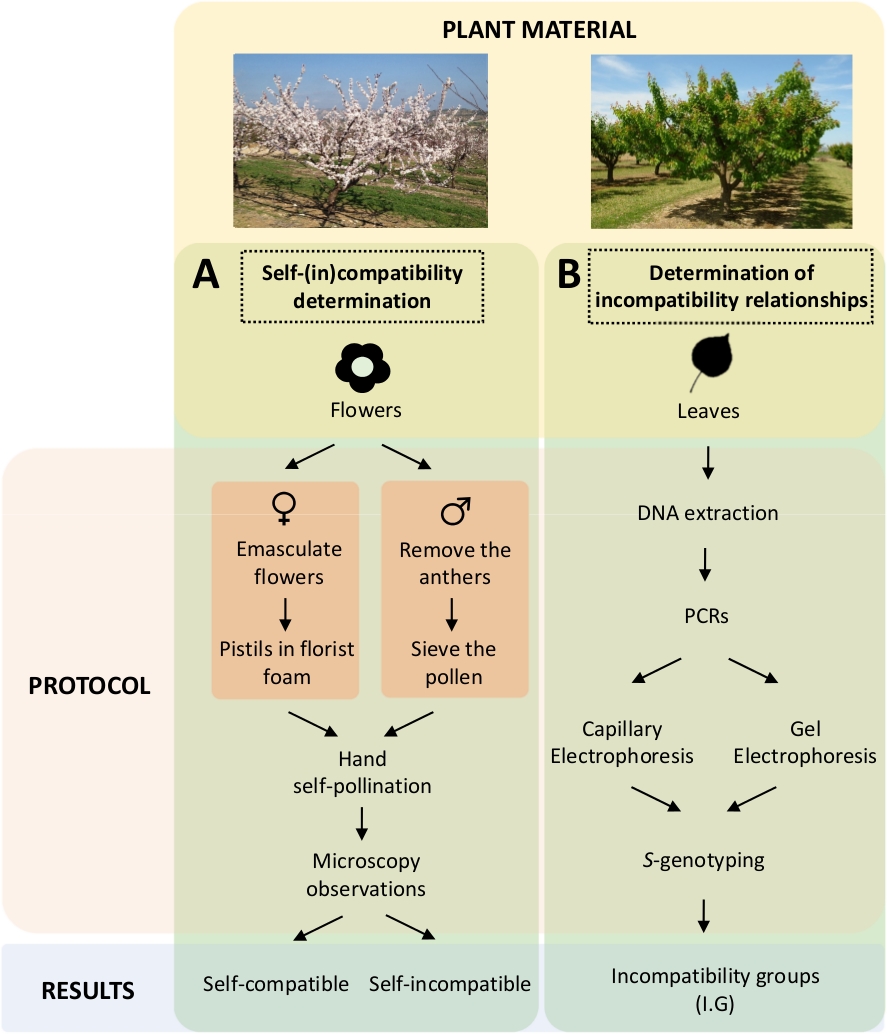
Denne fremgangsmåde kræver, at hver sorts kompatibilitet bestemmes ved kontrollerede selv- og krydsbestøvninger i laboratoriet (figur 5A) samtidig med karakteriseringen af S-genotypenved genetisk analyse ( figur5B). Som følge heraf kan bestøvningskravene for hver sort og uforenelighedsrelationerne mellem abrikoskulterne bestemmes.

Figur 1. Forsøgsforsøg, der er oprettet til bestemmelse af selv-in)kompatibilitet i abrikos.
(A) Blomster på ballonstadiet (sorte pile) i feltet. (B) Sigt af pollenkorn ved hjælp af en finmasket. cC) Pistils anbragt på blomsterskum i vand. (D) Håndbestøvning af støvveje ved hjælp af en pensel. Klik her for at se en større version af dette tal.

Figur 2. Diagrammatisk repræsentation af gametofytiske uforenelighed relationer i abrikos blomster.
(A) I Gametophytic Self-Incompatibility (GSI), både kompatible og uforenelige pollenkorn spire på stigmatisering. Pollenkornet bærer en af to S-alleleraf den oprindelige genotype, i dette tilfælde enten S1 eller S2. Hvis S-allelaf pollen korn matcher en af de to S-alleleraf pistil, i dette tilfælde S1S3, pollen rør vækst er hæmmet i den øverste tredjedel af stilen. ( B) Spiring af pollenkorn på stigmatiseringsoverfladen. (C) Pollen rør anholdt i stil angiver en uforenelig adfærd. (D) Pollen rør vokser langs stilen. (E) Pollenrør i bunden af den stil, der angiver en kompatibel adfærd. Vægtstænger, 100 μm. Klik her for at se en større version af dette tal.

Figur 3. Repræsentative resultater af pollenspiration og pollenrørvækst gennem stilen for selvkompatible og selv uforenelige sorter efter selvbestøvninger.
Procentdel af støvveje med pollenkorn spirende på stigmatisering overflade, med pollen rør i halvvejs stil, i bunden af den stil, og nå ovule. Klik her for at se en større version af dette tal.

Figur 4. PCR fragment forstærkning ved hjælp af fem primer par kombinationer til identifikation af S-alleler.
(A) Gene analysator output for SRc-(F / R) primere viser størrelsen af de to forstærkede fragmenter af RNase første intron region svarende til S-alleler. b) PCR-forstærkning ved hjælp af primere PruC2/PruC4R til identifikation af S6 og S9 alleler. cC) PCR-produkter, der er fremstillet ved hjælp af de specifikke primere SHLM1 og SHLM2 til differentiering af S1-allel og SHLM3 og SHLM4 for at skelne mellem S7-allelen. (D) PCR forstærkning med AprFBC8-(F/R) primere til identifikation af Sc og S8 alleler. MI:1 kb DNA Stige. MII: 100 bp DNA Stige. Klik her for at se en større version af dette tal.

Figur 5. Forsøgsdesignets skema til belysning af selv- og inter-(in) kompatibilitetsrelationer i abrikoskulter.
(A) Bestemmelse af selv-(in)kompatibilitet ved kontrollerede bestøvninger i laboratoriet. (B) Arbejdsgang for S-allelidentifikation ved molekylære tilgange. Klik her for at se en større version af dette tal.
| PCR-mastermix | Thermocycler betingelser | ||||||
| Komponenter | Endelig koncentration | 15 μL reaktion | Trin i cyklus | Temperatur | Tid | Cykler | |
| 10x NH4 Reaktionsbuffer | 10x | 1,5 μL | Indledende denaturering | 94 °C | 3 min. | 1 | |
| 50 mM MgCl2 opløsning | 25 mM | 1,2 μL | Denaturering | 94 °C | 1 min. | 35 | |
| 100 mM dNTP | 2,5 mM | 0,6 μL | Udglødning | 55 °C | 1 min. | ||
| Primer SRc-F | 10 μM | 0,6 μL | Udvidelse | 72 °C | 3 min. | ||
| Primer SRc-R | 10 μM | 0,6 μL | Endelig forlængelse | 72 °C | 5 min. | 1 | |
| 500 U Taq DNA Polymerase | 0,5 U | 0,2 μL | 4 °C | Holde | |||
| H2O | 8,3 μL | ||||||
| Komponenter | Endelig koncentration | 25 μL reaktion | Trin i cyklus | Temperatur | Tid | Cykler | |
| 10x PCR-buffer | 10x | 2,5 μL | Indledende denaturering | 94 °C | 2 min. | 1 | |
| 5x Q-løsning | 5x | 5 μL | Denaturering | 94 °C | 10 s | 10 | |
| 100 mM dNTP | 2,5 mM | 0,5 μL | Udglødning | 55 °C | 2 min. | ||
| Primer PruC2 | 10 μM | 0,2 μL | Udvidelse | 68 °C | 2 min. | ||
| Primer C4R | 10 μM | 0,2 μL | Denaturering | 94 °C | 10 s | 25 | |
| 250 U Taq DNA Polymerase | 10 Kr. | 0,13 μL | Udglødning | 58 °C | 2 min. | ||
| H2O | 15,5 μL | Udvidelse* | 68 °C | 2 min. | |||
| Endelig forlængelse | 72 °C | 5 min. | 1 | ||||
| 4 °C | Holde | ||||||
| * med 10 s tilføjet hver cyklus til 68 %C udvidelse trin. | |||||||
| Komponenter | Endelig koncentration | 25 μL reaktion | Trin i cyklus | Temperatur | Tid | Cykler | |
| 10x PCR-buffer | 10x | 2,5 μL | Indledende denaturering | 94 °C | 2 min. | 1 | |
| 5x Q-løsning | 5x | 5 μL | Denaturering | 94 °C | kr. | 35 | |
| 100 mM dNTP | 2,5 mM | 0,5 μL | Udglødning | 62 °C | 1,5 min. | ||
| Primer SHLM1 | 10 μM | 0,2 μL | Udvidelse | 72 °C | 2 min. | ||
| Primer SHLM2 | 10 μM | 0,2 μL | Endelig forlængelse | 72 °C | 5 min. | 1 | |
| 250 U Taq DNA Polymerase | 10 Kr. | 0,13 μL | 4 °C | Holde | |||
| H2O | 15,5 μL | ||||||
| Komponenter | Endelig koncentration | 20 μL reaktion | Trin i cyklus | Temperatur | Tid | Cykler | |
| 5x PCR-buffer | 5x | 4 μL | Indledende denaturering | 98 °C | kr. | 1 | |
| dNTP | 2,5 mM | 1,6 μL | Denaturering | 98 °C | 10 s | 35 | |
| Primer SHLM3 | 10 μM | 1 μL | Udglødning | 51 °C | kr. | ||
| Primer SHLM4 | 10 μM | 1 μL | Udvidelse | 72 °C | 1 min. | ||
| 100 U DNA Polymerase | 5 U | 0,2 μL | Endelig forlængelse | 72 °C | 5 min. | 1 | |
| H2O | 12,4 μL | 4 °C | Holde | ||||
| Komponenter | Endelig koncentration | 25 μL reaktion | Trin i cyklus | Temperatur | Tid | Cykler | |
| 10x PCR-buffer | 10x | 2,5 μL | Indledende denaturering | 94 °C | 2 min. | 1 | |
| 100 mM dNTP | 2,5 mM | 2 μL | Denaturering | 94 °C | kr. | 35 | |
| Primer FBC8-F | 10 μM | 1 μL | Udglødning | 55 °C | 1,5 min. | ||
| Primer FBC8-R | 10 μM | 1 μL | Udvidelse | 72 °C | 2 min. | ||
| 250 U Taq DNA Polymerase | 10 Kr. | 0,125 μL | Endelig forlængelse | 72 °C | 5 min. | 1 | |
| H2O | 17,4 μL | 4 °C | Holde | ||||
Tabel 1. Reaktions- og cykelforhold for forskellige primerkombinationer, der anvendes i denne protokol.
| Primere | Sekvens | Reference |
| SRc-F | 5'-CTCGCTTTTGTTGTTCTTGC-3' | 18 |
| SRc-R | 5'-GGCCATTGTTGCACCCCTTG-3' | 18 |
| Pru-C2 | 5'-CTTTGGCCAAGAGTAATTATTCAAACC-3' | 35 |
| Pru-C4R | 5'-GGATGTGGTACGATTGAAGCG-3' | 35 |
| SHLM1-F | 5'-GGTGGAGGTGATAAGGTAGCC-3' | 17 |
| SHLM2-R | 5'-GGCTGCATAAGGAAGCTGTAGG-3' | 17 |
| SHLM3-F | 5'-TATATCTTACTCTTTTGGC-3' | 17 |
| SHLM4-R | 5'-CACTATGATAATGTATG-3' | 17 |
| AprFBC8-F | 5'-CATGGAAAAAGCTGACTTATGG-3' | 26 |
| AprFBC8-R | 5'-GCCTCTAATGTCATCTACTCTTAG-3' | 26 |
Tabel 2. Primere, der anvendes i denne protokol, sekvens og reference for S-genotype karakterisering i Prunus armenienca.
| Sort | SRc-(F/R) (bp) | PruC2/PruC4R (bp) | SHLM1/SHLM2 (bp) | SHLM3/SHLM4 (bp) | AprFBC8-(F/R) (bp) | S-Genotype | Inkompatibilitetsgruppe (I.G) |
| Wonder Barneseng8 | 420, 420 | 749, 1386 | S6S9 | Viii | |||
| Magisk barneseng8 | 334, 420 | 749 | S2S9 | Xx | |||
| Guldstrike8 | 334, 420 | 749 | S2S9 | - | |||
| T06917 | 334, 408 | 650 | S1S2 | Ⅰ | |||
| T12017 | 334, 408 | 650 | S1S2 | - | |||
| C-6 | 334, 408 | 413 | S2S7 | Iv | |||
| Cooper Cot8 | 274, 408 | 650 | S1S3 | Xviii | |||
| Apriqueen (Apriqueen) | 358, 358 | 500 | ScSc | - | |||
| Bergecot 8 delteet år. | 334, 358 | 500 | S2Sc | - | |||
| Forår Blush8 | 274, 358 | 150 | S3S8 | Xxi |
Tabel 3. S-genotyping af abrikoskulter med fem primerpar, der anvendes i denne protokol og inkompatibilitetsgruppetildeling. S De forskellige polymerasekædereaktionsproduktstørrelser for S-alleler, der er forstærket ved hjælp af SRc-(F/R), PruC2/PruC4R, SHLM1/ SHLM2, SHLM3/SHLM4 og AprFBC8-(F/R) primere, er vist i tabellen. S
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Traditionelt var de fleste kommercielle abrikoseuropæiske sorter selvkompatible36. Ikke desto mindre har brugen af nordamerikanske selv uforenelige sorter som forældre i avlsprogrammer i de seneste årtier resulteret i frigivelsen af et stigende antal nye selv uforenelige sorter med ukendte bestøvningskrav7,8,37. Det bliver således stadig vigtigere at bestemme selv- og indbyrdes kompatibilitetsforhold hos abrikoskulter. Dette forstærkes i de områder, hvor vinteren nedkøling er faldende, da høje år til år variationer i tidspunktet for blomstring forhindrer sammenfald i blomstring af sorter og deres pollenizere i mange tilfælde, især i sorter med høj nedkøling krav38. Den metode, der er beskrevet heri, kombinere håndbestøvning, mikroskopi og genetiske analyser har været meget nyttigt at bestemme selv (i) kompatibilitet af hver sort og til at etablere dens potentielle bestøver sorter.
Bestøvningskrav kan bestemmes ved hjælp af feltkontrolforsøg under plantageforhold11,39. Men, redegørelsen til eksterne faktorer, herunder meteorologiske ugunstige forhold kan forårsage bestøvning fiasko10, hvilket kan resultere i fejlagtige diagnoser af selv-uforenelighed. Den metode, der er beskrevet heri, gør det muligt at evaluere selv-(in)kompatibilitet mere præcist ved mikroskopi observationer af håndbestøvede blomster i laboratorie-kontrollerede forhold, undgå miljøpåvirkning. Desuden er denne fremgangsmåde gør det muligt at analysere et højere antal sorter om året, da kun et lille antal blomster er påkrævet i stedet for flere voksne træer for hver sort, der kræves i feltforsøg40.
Der kan etableres uforenelighedsrelationer , der kombinerer håndbestøvninger og mikroskopi14. Bestøvningerne kan dog kun udføres i en kort periode i blomstringssæsonen om foråret, og voksne træer i nærheden af laboratoriet er nødvendige, da levetiden for de indsamlede blomster er meget kort. Således er antallet af uforenelighed relationer, der kan analyseres ved kontrollerede hånd-bestøvninger i hver sæson er meget lav. Karakteriseringen af de gener, der er kodet af S-locus, har gjort det muligt at udvikle PCR-baserede metoder til S-allelgenotyping18,41. Denne fremgangsmåde Sfremskynder S-allel identifikation, da det ikke kræver blomster, og forsøgene kan udføres med enhver vegetativt væv42. Dette forlænger den periode, hvor plantemateriale, normalt unge blade, kan indsamles43. Desuden kan bladene være frysetørret eller frosset, således at analysen kan gøres på ethvert tidspunkt af året, i modsætning til bestøvninger, der kun kan gøres på friske blomster i blomstring sæson44. En yderligere fordel er, at blade kan indsamles fra unge træer, selv før de kommer ind blomstring alder, lette indsamlingen af prøver og tidlig opnåelse af resultater45.
Den genetiske analyse muliggør en bedre differentiering af selv-uforenelighedsredler , da den giver præcise resultater af forstærkede fragmentstørrelser21,46. Hidtil er 33 S-allelerblevet identificeret i abrikos12,,18,20,21,22,23,24, som har gjort det muligt at etablere 36 uforenelighedsgrupper baseret på S-genotype8,9,17,25,26,27. På den anden side er en ulempe ved denne metode, at forskellige alleler i samme områdestørrelse eller mutationer fejlagtigt kan identificeres som den samme allel. Således Sc og S8 alleler er identiske for RNase sekvens, men en 358-bp indsættelse findes i SFB genet af Sc19. Ligeledes er den første intron region af alleles S1 og S7 identiske og kan ikke skelnes ved hjælp af primere SRc-F/SRc-R. Desuden er der fundet flere homologier, såsom S6 og S528 eller S20 og S55og S7, S13 (EF062341) og S4617, fordi nogle af disse alleler er blevet delvist sekventeret eller af fejl under PCR-forstærkning, og der derfor er behov for yderligere arbejde for at skelne dem korrekt.
PCR-analyse og S-RNase-sekvensering er tilstrækkelige til at etablere uforenelighedsrelationer ved at identificere S-allelerog fordelingen af sorter i deres tilsvarende uforenelighedsgruppe8,17,26og27 . Denne metode har imidlertid den begrænsning, at den forhindrer, at der fastlægges selv-(in)kompatibilitet for bestemte abrikoskulter. Selvkompatibilitet (SC) har været forbundet med særlige S-alleleri andre Prunus arter47, som mandel (Sf)48,49 eller søde kirsebær (S4')50,51. Men i abrikos, Sc allel, som har været forbundet med SC21, kan fejlagtigt identificeres som S8, en selvkompatibel allel19,22, og mulige mutationer ikke er knyttet til S locus, som M-locus12,52,giver SC er blevet identificeret. For nylig Mer M-locus blevet genotyped ved hjælp af SSR markører12. Derfor har den genetiske identifikation af SC for abrikos genotyper brug for yderligere forskning, og for at undgå fejl på grund af faktorer, der ikke er knyttet til S locus, i dette arbejde karakterisering af selv-(i)kompatibilitet er blevet bestemt også ved fænotypebestemmelse adfærd pollen rør gennem støvstempling af selvbestøvede blomster.
Den metode, der er beskrevet heri, og som kombinerer bestemmelse af selv-(in)kompatibilitet ved håndbestøvning under laboratorieforhold med den efterfølgende observation af pollenrørenes Sopførsel i støvlen af kontrollerede selvbestøvninger under fluorescensmikroskopien, og pcr-analysens identifikation af S-genotype gør det muligt at fastsætte bestøvningskravene for abrikoskultivare. Dette giver en værdifuld information til avlere og opdrættere, da det gør det muligt at etablere uforenelighed forholdet mellem sorter til at vælge egnede bestøvere i udformningen af nye plantager samt at vælge passende forældre til at designe nye kors i abrikos avlsprogrammer.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne har intet at afsløre.
Acknowledgments
Denne forskning blev finansieret af Ministerio de Ciencia, Innovación y Universidades-European Regional Development Fund, Den Europæiske Union (AGL2016-77267-R og AGL2015-74071-JIN); Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (RFP2015-00015-00, RTA2017-00003-00); Den Europæiske Socialfond for Gobierno de Aragón, Den Europæiske Union (Grupo Consolidado A12_17R), Fundación Biodiversidad og Agroseguro S.A.
Materials
| Name | Company | Catalog Number | Comments |
| Agarose D1 Low EEO | Conda | 8010.22 | |
| BIOTAQ DNA Polymerase kit | Bioline | BIO-21060 | |
| Bright field microscope | Leica Microsystems | DM2500 | |
| CEQ System Software | Beckman Coulter | ||
| DNeasy Plant Mini Kit | QIAGEN | 69106 | |
| dNTP Set, 4 x 25 µmol | Bioline | BIO-39025 | |
| GenomeLab DNA Size Standard Kit - 400 | Beckman Coulter | 608098 | |
| GenomeLab GeXP Genetic Analysis System | Beckman Coulter | ||
| GenomeLab Separation Buffer | Beckman Coulter | 608012 | |
| GenomeLab Separation Gel LPA-1 | Beckman Coulter | 391438 | |
| HyperLadder 100bp | Bioline | BIO-33029 | |
| HyperLadder 1kb | Bioline | BIO-33025 | |
| Image Analysis System | Leica Microsystems | ||
| Molecular Imager VersaDoc MP 4000 system | Bio-Rad | 170-8640 | |
| NanoDrop One Spectrophotometer | Thermo Fisher Scientific | 13-400-518 | |
| pH-Meter BASIC 20 | Crison | ||
| Phusion High-Fidelity PCR Kit | Thermo Fisher Scientific | F553S | |
| Power Pack P 25 T | Biometra | ||
| Primer Forward | Isogen Life Science | ||
| Primer Reverse | Isogen Life Science | ||
| Quantity One Software | Bio-Rad | ||
| Stereoscopic microscope | Leica Microsystems | MZ-16 | |
| Sub-Cell GT | Bio-Rad | ||
| SYBR Safe DNA Gel Stain | Thermo Fisher Scientific | S33102 | |
| T100 Thermal Cycler | Bio-Rad | 1861096 | |
| Taq DNA Polymerase | QIAGEN | 201203 | |
| Vertical Stand Autoclave | JP Selecta |
References
- Silva, N. F., Goring, D. R.
- Charlesworth, D., Vekemans, X., Castric, V., Glémin, S. Plant self-incompatibility systems: A molecular evolutionary perspective. New phytologist. 168, 61-69 (2005).
- Tao, R., et al. Identification of stylar RNases associated with gametophytic self-incompatibility in almond (Prunus dulcis). Plant and Cell Physiology. 38, 304-311 (1997).
- Ushijima, K., et al. Structural and transcriptional analysis of the self-incompatibility locus of almond: Identification of a pollen-expressed F-box gene with haplotype-specific polymorphism. The Plant cell. 15, 771-781 (2003).
- Bedinger, P. A., Broz, A. K., Tovar-Mendez, A., McClure, B. Pollen-Pistil Interactions and Their Role in Mate Selection. Plant Physiology. 173, 79-90 (2017).
- Guerra, M. E., Rodrigo, J. Japanese plum pollination: A review. Scientia Horticulturae. 197, 674-686 (2015).
- Zhebentyayeva, T., Ledbetter, C., Burgos, L., Llacer, G. Apricot. Fruit Breeding. Badenes, M. L., Byrne, D. , Springer. Boston, MA. 415-458 (2012).
- Herrera, S., Lora, J., Hormaza, J. I., Herrero, M., Rodrigo, J. Optimizing Production in the New Generation of Apricot Cultivars: Self-incompatibility, S-RNase Allele Identification, and Incompatibility Group Assignment. Frontiers in Plant Science. 9, 527 (2018).
- Egea, J., Burgos, L. Detecting Cross-incompatibility of Three North American Apricot Cultivars and Establishing the First Incompatibility Group in Apricot. Journal of the American Society for Horticultural Science. 121, 1002-1005 (1996).
- Rodrigo, J., Herrero, M. Effects of pre-blossom temperatures on flower development and fruit set in apricot. Scientia Horticulturae. 92, 125-135 (2002).
- Julian, C., Herrero, M., Rodrigo, J. Flower bud differentiation and development in fruiting and non-fruiting shoots in relation to fruit set in apricot (Prunus armeniaca). Trees. 24, 833-841 (2010).
- Muñoz-Sanz, J. V., Zuriaga, E., López, I., Badenes, M. L., Romero, C. Self-(in)compatibility in apricot germplasm is controlled by two major loci, S and M. BMC Plant Biology. 17, 82 (2017).
- Burgos, L., Berenguer, T., Egea, J. Self- and Cross-compatibility among Apricot Cultivars. HortScience. 28, 148-150 (1993).
- Rodrigo, J., Herrero, M. Evaluation of pollination as the cause of erratic fruit set in apricot "Moniqui". Journal of Horticultural Science. 71, 801-805 (1996).
- Milatović, D., Nikolić, D., Krška, B. Testing of self-(in)compatibility in apricot cultivars from European breeding programmes. Horticultural Science. 40 (2), 65-71 (2013).
- Milatović, D., Nikolić, D., Fotirić-Aksić, M., Radović, A. Testing of self-(in)compatibility in apricot cultivars using fluorescence microscopy. Acta Scientiarum Polonorum, Hortorum Cultus. 12 (6), 103-113 (2013).
- Herrera, S., Rodrigo, J., Hormaza, J. I., Lora, J. Identification of Self-Incompatibility Alleles by Specific PCR Analysis and S-RNase Sequencing in Apricot. Int J Mol Sci. 19, 3612 (2018).
- Romero, C., et al. Analysis of the S-locus structure in Prunus armeniaca L. Identification of S-haplotype specific S-RNase and F-box genes. Plant Molecular Biology. 56, 145-157 (2004).
- Halász, J., Pedryc, A., Hegedus, A. Origin and dissemination of the pollen-part mutated SC haplotype which confers self-compatibility in apricot (Prunus armeniaca). New Phytologist. 176, 792-803 (2007).
- Halász, J., Hegedus, A., Hermán, R., Stefanovits-Bányai, É, Pedryc, A. New self-incompatibility alleles in apricot (Prunus armeniaca L.) revealed by stylar ribonuclease assay and S-PCR analysis. Euphytica. 145, 57-66 (2005).
- Vilanova, S., Romero, C., Llacer, G., Badenes, M. L., Burgos, L. Identification of Self-(in)compatibility Alleles in Apricot by PCR and Sequence Analysis. Journal of the American Society for Horticultural Science. 130, 893-898 (2005).
- Feng, J., et al. Detection and transcript expression of S-RNase gene associated with self-incompatibility in apricot (Prunus armeniaca L.). Molecular Biology Reports. 33, 215-221 (2006).
- Zhang, L., et al. Identification of self-incompatibility (S-) genotypes of Chinese apricot cultivars. Euphytica. 160, 241-248 (2008).
- Wu, J., et al. Identification of S-haplotype-specific S-RNase and SFB alleles in native Chinese apricot (Prunus armeniaca L). Journal of Horticultural Science and Biotechnology. 84, 645-652 (2009).
- Szabó, Z., Nyéki, J. Blossoming, fructification and combination of apricot varieties. Acta Horticulturae. 293, 295-302 (1991).
- Halász, J., Pedryc, A., Ercisli, S., Yilmaz, K. U., Hegedűs, A. S-genotyping supports the genetic relationships between Turkish and Hungarian apricot germplasm. Journal of the American Society for Horticultural Science. 135, 410-417 (2010).
- Lachkar, A., et al. Identification of self-(in)compatibility S-alleles and new cross-incompatibility groups in Tunisian apricot (Prunus armeniaca L.) cultivars. The Journal of Horticultural Science and Biotechnology. 88, 497-501 (2013).
- Pérez-Pastor, A., Ruiz-Sánchez, M. C., Domingo, R., Torrecillas, A. Growth and phenological stages of Búlida apricot trees in South-East. Agronomie. 24, 93-100 (2004).
- Williams, J. H., Friedman, W. E., Arnold, M. L. Developmental selection within the angiosperm style: using gamete DNA to visualize interspecific pollen competition. Proceedings of the National Academy of Sciences of the United States of America. 96, 9201-9206 (1999).
- Julian, C., Herrero, M., Rodrigo, J. Anther meiosis time is related to winter cold temperatures in apricot (Prunus armeniaca L.). Environmental and Experimental Botany. 100, 20-25 (2014).
- Guerra, M. E., López-Corrales, M., Wünsch, A., Rodrigo, J. Lack of Fruit Set Caused by Ovule Degeneration in Japanese Plum. Journal of the American Society for Horticultural Science. 136 (6), 375-381 (2011).
- Guerra, M. E., Wünsch, A., López-Corrales, M., Rodrigo, J. Flower Emasculation as the Cause for Lack of Fruit Set in Japanese Plum Crosses. Journal of the American Society for Horticultural Science. 135 (6), 556-562 (2010).
- Hormaza, J. I., Pinney, K., Polito, V. S. Correlation in the tolerance to ozone between sporophytes and male gametophytes of several fruit and nut tree species (Rosaceae). Sexual Plant Reproduction. 9, 44-48 (1996).
- Alcaraz, M. L., Hormaza, J. I., Rodrigo, J. Pistil Starch Reserves at Anthesis Correlate with Final Flower Fate in Avocado (Persea americana). PLOS ONE. 8 (10), 78467 (2013).
- Tao, R., et al. Molecular typing of S-alleles through Identification, Characterization and cDNA cloning for S-RNases in Sweet Cherry. Journal of the American Society for Horticultural Science. 124, 224-233 (1999).
- Burgos, L., et al. The self-compatibility trait of the main apricot cultivars and new selections from breeding programmes. Journal of Horticultural Science. 72, 147-154 (1997).
- Hormaza, J. I., Yamane, H., Rodrigo, J. Apricot. Genome Mapping and Molecular Breeding in Plants, Volume 4 Fruits and Nuts. Kole, C. , Springer. New York, NY. 171-187 (2007).
- Benmoussa, H., Ghrab, M., Ben Mimoun, M., Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agricultural and Forest Meteorology. 239, 34-46 (2017).
- Rodrigo, J., Herrero, M., Hormaza, J. I. Pistil traits and flower fate in apricot (Prunus armeniaca). Annals of Applied Biology. 154, 365-375 (2009).
- Williams, R. R. Techniques used in fruit-set experiments. Towards Regulated Cropping. Williams, R. R., Wilson, D. , Grower Books. London. 57-61 (1970).
- Sutherland, B. G., Robbins, T. P., Tobutt, K. R. Primers amplifying a range of Prunus S-alleles. Plant Breeding. 123, 582-584 (2004).
- Murray, M. G., Thompson, W. F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Research. 8, 4321-4325 (1980).
- Porebski, S., Bailey, L. G., Baum, B. R. Modification of a CTAB DNA Extraction Protocol for Plants Containing High Polysaccharide and Polyphenol Components. Plant Molecular Biology Reporter. 15 (1), 8-15 (1997).
- Rogers, S. O., Bendich, A. J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Molecular Biology. 5 (2), 69-76 (1985).
- Hormaza, J. I. Molecular characterization and similarity relationships among apricot (Prunus armeniaca L.) genotypes using simple sequence repeats. Theoretical and Applied Genetics. 104, 321-328 (2002).
- Sonneveld, T., Tobutt, K. R., Robbins, T. P. Allele-specific PCR detection of sweet cherry self-incompatibility (S) alleles S1 to S16 using consensus and allele-specific primers. Theoretical and Applied Genetics. 107, 1059-1070 (2003).
- Hegedus, A., Lénárt, J., Halász, J. Sexual incompatibility in Rosaceae fruit tree species: molecular interactions and evolutionary dynamics. Biologia Plantarum. 56 (2), 201-209 (2012).
- Fernández i Martí, A., Gradziel, T. M., Socias i Company, R. Methylation of the Sf locus in almond is associated with S-RNase loss of function. Plant Molecular Biology. 86, 681-689 (2014).
- Company, R. S. i, Kodad, O., Martí, A. F. i, Alonso, J. M. Mutations conferring self-compatibility in Prunus species: From deletions and insertions to epigenetic alterations. Scientia Horticulturae. 192, 125-131 (2015).
- Boskovic, R., Tobutt, K. R. Correlation of stylar ribonuclease zymograms with incompatibility alleles in sweet cherry. Euphytica. 90, 245-250 (1996).
- Cachi, A. M., Wünsch, A. S-genotyping of sweet cherry varieties from Spain and S-locus diversity in Europe. Euphytica. 197 (2), 229-236 (2014).
- Zuriaga, E., et al. An S-locus Independent Pollen Factor Confers Self-Compatibility in "Katy" Apricot. PLoS ONE. 8 (1), 53947 (2013).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}