As shown in Figure 2A, CAF diet feeding produces a 2.5-fold increase in energy intake relative to chow controls, based on data from three cohorts of male Sprague Dawley rats, that is consistent over 6 weeks. Other studies have confirmed that this extent of hyperphagia is sustained over 1021 and 1622 week experiments. The weight curve (Figure 2B) indicates CAF diet feeding leads to a 20% difference in mean body weight compared with control after 3–4 weeks of diet, comparable to the body weight gain consistent with the onset of obesity in humans. At 6 weeks, mean difference in body weight gain between CAF and control groups is 67% (Figure 2C) and adiposity, determined by EchoMRI, is approximately doubled (Figure 2D). Cafeteria-fed rats typically eat 5–10% of their daily energy as chow (~5 g/rat/day).
Macronutrient intake profiles can be calculated on food intake measurement days using nutritional information from the product manufacturers. We observe consistent macronutrient intakes across sexes and ages, with CAF-fed rats consuming approximately 8% of energy as protein, 34% as fat and 58% as carbohydrate. Our maintenance chow provides 22% protein, 13% fat and 65% carbohydrate. Relative to estimates of macronutrient consumption in human populations (18.3% protein, 44.9% carbohydrate and 30.9% fat in Australia23; 15.7% protein, 48.7% carbohydrate and 33.7% fat in the United States of America24), our CAF-fed rats consume a lower proportion of energy as protein, a higher proportion as carbohydrate, and a comparable proportion from fat. However, when considering absolute intake, CAF-fed rats overeat all three macronutrients relative to controls (Figure 3A–C for males, Figure 3D–F for females), indicating that they are not protein deficient. Rather, the macronutrient composition results from the dramatic hyperphagia evoked, which is driven predominantly by excess carbohydrate and fat intake, not usually observed in human subjects who tend to become overweight and develop obesity more gradually.
A recent study we conducted suggests that female rats may be particularly vulnerable to the obesogenic effects of CAF diet. Energy intake was 3.8 times greater in CAF-fed females relative to controls, which was sustained over 6 weeks (Figure 4A). A 20% weight difference between groups was observed after only 2 weeks of CAF exposure (Figure 4B). After 6 weeks of diet exposure, body weight gain was 12 times greater in CAF rats (Figure 4C) and fat mass was doubled (Figure 4D) compared with healthy controls. The suggestion of greater susceptibility to diet-induced weight gain in females is supported by an earlier study by Sclafani and Gorman, which showed that a cafeteria diet induced significantly greater weight gain in females than in males25.
Providing too little of each CAF food may artificially constrain measurements of energy intake. This is most easily addressed by checking that there is some food remaining in the CAF cages 24 h after feeding (when the food is refreshed). Figure 5 shows how the use of multiple food sets for food intake can lead to substantial shifts in macronutrient intake despite comparable overall energy intake. In this instance, the variability was due primarily to the balance of fat and carbohydrate against a backdrop of consistent protein intake. Analyses of the food sets used indicated that high carbohydrate intakes were observed when a highly-preferred cake was provided; higher fat intakes were observed when this cake was absent.

Figure 1: Example of cafeteria diet across three days. CAF diet should consist of varied food each day to induce sustained hyperphagia. Preparing CAF foods can be streamlined by placing each cage’s food into a designated container. This allows for easy, well-timed delivery into each cage. Day 1 = chicken nuggets, beef-flavored dog food, chocolate cream biscuits, jam roll, high-fat purified diet. Day 2 = meat pie, chicken nuggets, scotch finger biscuits, caramel mud cake. Day 3 = dim sum, chicken-flavored dog food, custard cream biscuits, blueberry cheesecake. Please click here to view a larger version of this figure.
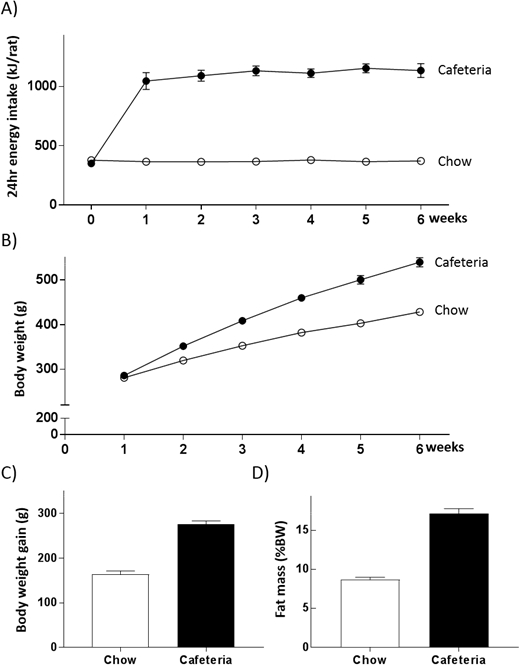
Figure 2: Representative results in multiple male cohorts. CAF diet produces a consistent increase in energy intake (A), and body weight (B) over 6 weeks in male adult rats. This is accompanied by significant increases in body weight gain (C) and fat mass (D) when assessed by EchoMRI at 4 weeks of diet. Data are represented as mean ± SEM; n = 48 for individual data; n = 12 for energy intake data (cage as the unit of analysis). Please click here to view a larger version of this figure.
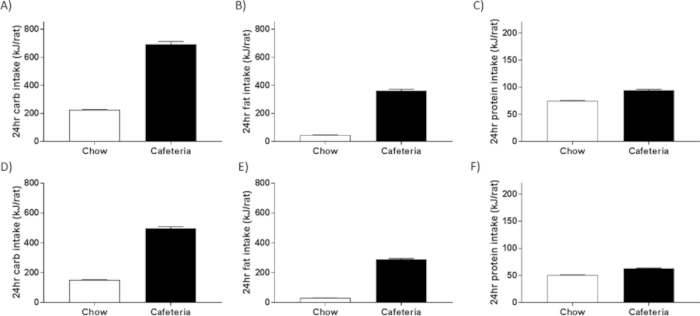
Figure 3: Macronutrient intake in multiple male cohorts. CAF diet exposure increases total intake of carbohydrate (A), fat (B) and protein (C) for male rats. These increases are comparable in female rats for carbohydrate (D), fat (E) and protein (F) intakes. Data are represented as mean ± SEM; n = 12 or n = 4 cages (males and females respectively) averaged over 6 weeks. Please click here to view a larger version of this figure.
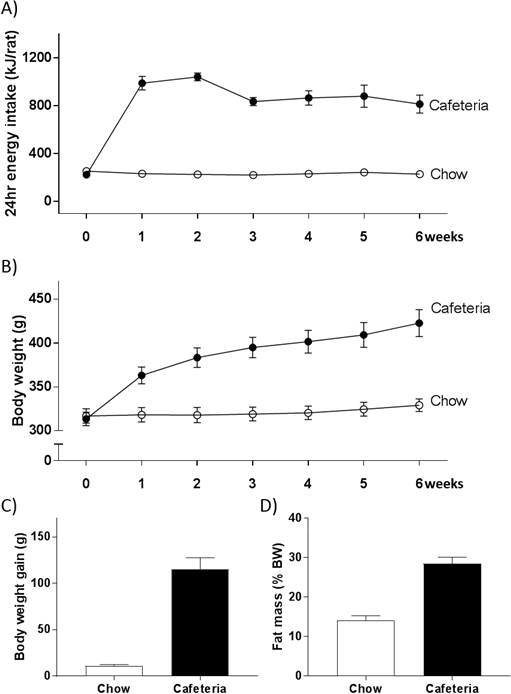
Figure 4: Representative results in a female cohort. Cafeteria diet produces a consistent increase in energy intake (A) and body weight (B) over 6 weeks in female adult rats. After 6 weeks of diet a substantial difference in body weight gain (C) and fat mass (D) is also observed. Data are represented as mean ± SEM; n = 12 for individual data; n = 4 for energy intake data. Please click here to view a larger version of this figure.
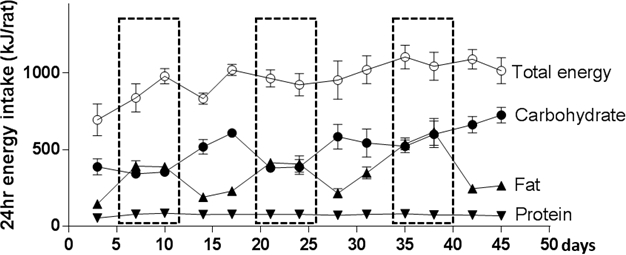
Figure 5: Macronutrient intake over time. Using multiple food sets to assess food intake can lead to differences in macronutrient intake over time given differences in individual preferences in a cohort of male rats. Data are represented as mean ± SEM; n = 4 cages. Note that overall macronutrient intake matches the pattern usually observed; CAF-fed rats consumed 60% energy as carbohydrates, 33% fat and 8% protein, as energy. Please click here to view a larger version of this figure.
| Monday | Tuesday | Wednesday | Thursday | Friday | Saturday | Sunday | |
| DAILY FOODS | Healthy chow and potable water | ||||||
| CAKE | A | D | B | A | C | B | E |
| PROTEIN 1 | A | C | D | A | C | D | A |
| PROTEIN 2 | B | B | A | B | B | A | C |
| COOKIE | B | A | C | B | D | C | A |
| OPTIONAL ADDITIONAL DAILY FOODS | HFHS chow, 10% sucrose solution | ||||||
Table 1: Example weekly food plan for the cafeteria diet. The CAF diet promotes hyperphagia by providing a variety of palatable foods that are varied daily, as shown in this example weekly food plan. Letters A-E denote unique foods for that food group (for example, Cake A might denote chocolate mud cake and Cake B vanilla sponge). Food intake days, shaded in grey, should be positioned evenly across the week and ideally are kept as consistent as possible. While CAF diet always includes continuous access to healthy chow and water, optional additional daily foods can include HF or HFHS chows, and 10% sucrose solution.
| PER 100g (derived from manufacturer) | ENERGY PER 1g | Recommended starting value (kJ/rat) | |||||||||||
| Food | Energy (kJ) | Protein (g) | Total Fat (g) | Saturated Fat (g) | Total Carbohydrate (g) | Sugar (g) | Energy (kJ/g) | Protein (kJ) | Total Fat (kJ) | Saturated Fat (kJ) | Total Carbohydrate (kJ) | Sugar (kJ) | |
| Protein A | 830.00 | 6.00 | 6.10 | 3.20 | 28.90 | 2.20 | 8.19 | 1.02 | 2.26 | 1.18 | 4.91 | 0.37 | 350.00 |
| Protein B | 906.00 | 7.30 | 11.10 | 4.60 | 21.10 | 1.80 | 8.94 | 1.24 | 4.11 | 1.70 | 3.59 | 0.31 | 350.00 |
| Cake A | 1470.00 | 4.60 | 13.30 | 3.70 | 52.40 | 33.10 | 14.61 | 0.78 | 4.92 | 1.37 | 8.91 | 5.63 | 200.00 |
| Cake B | 1660.00 | 4.00 | 18.40 | 4.30 | 53.60 | 36.30 | 16.60 | 0.68 | 6.81 | 1.59 | 9.11 | 6.17 | 200.00 |
| Cookie A | 1920.00 | 4.30 | 20.60 | 12.70 | 63.20 | 33.20 | 19.10 | 0.73 | 7.62 | 4.70 | 10.74 | 5.64 | 200.00 |
| Cookie B | 2040.00 | 5.70 | 21.00 | 11.20 | 8.50 | 4.10 | 10.18 | 0.97 | 7.77 | 4.14 | 1.45 | 0.70 | 200.00 |
Table 2: Nutritional information for selected cafeteria food items. This table depicts the nutritional information obtained for several core items in the CAF diet. It is important to ensure that the daily options provide similar macronutrient availabilities, and that rats have access to adequate protein. For each daily set of foods used, it is helpful to calculate the overall energy density and the macronutrient content. The final column contains the recommended starting volume of each food (as energy per rat) for male Sprague Dawley rats at 200 g.
| CAGE | WATER (g) | CHOW (g) | Cake A (g) | Protein A (g) | Protein B (g) | Cookie B (g) | ||||||
| IN | OUT | IN | OUT | IN | OUT | IN | OUT | IN | OUT | IN | OUT | |
| 1 (CHOW) | ||||||||||||
| 2 (CAF) | ||||||||||||
| 3 (CHOW) | ||||||||||||
| 4 (CAF) | ||||||||||||
Table 3: Example food intake sheet. The weights of each CAF food item (g, per cage) should be carefully recorded on a spreadsheet.
