אנו מתארים כולל השתקפות פנימית מספר ובהירות (TIRF-N & B) גישה להדמיה לקביעת מצב oligomeric הממוצע של מולקולות הקולטן בקרום פלזמה של תאים חיים, מכוון בקישור הרכבה הקולטן דינמיקה לתפקוד הביולוגי של החלבונים (איור 1).
עם ליגתאי ומחייב, קולטנים ליזום את האות תאיים התמרה האותות בהתאם הקונפורמציה שלהם, oligomerization, שיתוף הפוטנציאלי קולטנים והרכב קרום. למרות החשיבות והייתה של קולטן oligomerization, מוכר כאירוע מפתח באיתות סלולרי1,2,3,4,5,6, 7, כמה שיטות יכול לזהות אירועים באשכולות ולמדוד את מידת הניסויים באשכולות8,9. כרך קונפוקלית וקד (x, y ≈ 300 ננומטר, z ≈ 900 nm) הוא פתר מספיק להוכחת אינטראקציה מולקולרית וסטויכמטריה, גם לאחר אופטימיזציה על ידי אלגוריתמים שיקום התמונה10. הרכב התת-יחידתי של חלבון oligomers אינו ניתן לפענוח על בסיס מרחבי גרידא גם על ידי שיטות ברזולוציה גבוהה ב-x, y ברזולוציה של 20-70 ננומטר כגון PALM11, סטורם12, ו ההיסטד13. יתר על כן, הרזולוציה הטמפורלית שלהם (בסדר של דקות לכל תמונה) לא יכול לעקוב אחר הקינטיקה בטווח של שניות. מולקולה בודדת צעד הלבנת פותרת את סטויכיומטריה של oligomers חלבון רק אם הם משותק14.
אחת השיטות הרב-תכליתיות ביותר כדי למדוד צפיפות ו oligomerization של חלבונים מתויגים פלואורוסקופים בתוך תמונות בודדות היא התפלגות בעוצמה מרחבית ניתוח (SpIDA), אשר נשענת על דגימה מרחבית. הוא חל על תאים כימיים קבועים ולחיות, ומאפשר ניתוח של מספר אזורים של העניין של התא בו זמנית באמצעות מיקרוסקופ קרינה רגילה15. לחילופין, שיטות רגע, כגון ספקטרוסקופיית מתאם פלואורסצנטית (fcs)16, היסטוגרמה לספירת פוטון (PCH)17, ומספר ובהירות (N & B)18,19, מתאימים לאולייומר כמותית דידות. שיטות אלה לנתח תנודות בעוצמה פלואורסצנטית כי ניתן להבחין בזמן כאשר fluorophores לפזר והחוצה של נפח התאורה. המוני התנודות באינטנסיביות ניתן לתאר באופן ייחודי על-ידי הבהירות המולקולרית של הזוהר (ε) והמספר הממוצע של fluorophores (n) בתוך כרך התאורה17 (איור 2). בדרך כלל, מקדם דיפוזיה של fluorophores ואת המספר הממוצע של מולקולות (הקשורים באופן הפוך לערך G (0) בתוך עוצמת התאורה ניתן להשיג על ידי FCS20. עם זאת, מאז זמן הדיפוזיה רק עם השורש העוקב של המסה, FCS אינו רגיש מספיק כדי לזהות שינויים במסה מולקולרית21. בפועל, לצבע אחד FCS אין אפשרות לזהות הדיסמזציה של קולטני הממברנה. PCH פותר תערובות של oligomers שונים במדויק. באמצעות יותר משני רגעים של התפלגות משרעת, הוא מזהה מולקולות של בהירות שונה התופסים את אותו נפח תאורה. סריקת FCS22 ופיתוחים, כגון מתאם זוג מעניין של בהירות מולקולרית (pcomb) גישה23, הציג כדי להאריך את מגוון הישימות של שיטות מתאם פלואורסצנטית במערכות ביולוגיות24 , השארו שיטות חד שונות חסרות יכולת של מדידות מהירות בשטח גדול של תא, הדורשות תצפיות רצופות רבות בכל פיקסל ורכישת נתונים בסדר השניות.
N & B הוא גרסה פשוטה של PCH הרואה רק את הרגעים הראשונים והשני של משרעת התפלגות הזריחה, כלומר את העוצמה ממוצע, , ואת השונות, σ2 (איור 2)18,19 ובגלל זה, זה לא יכול לקבוע את החלק הטוחנת של oligomers לא ידוע בתערובת, אבל רק מעריך את המצב הממוצע oligomeriטיזציה של התערובת. עם זאת, N & B יש את היתרון של עבודה עם סדרת זמן קטן יחסית של תמונות של תאים חיים מאשר PCH על בסיס פיקסל על ידי פיקסל, פשוט על ידי ניטור תנודות בזמן של עוצמת הקרינה. כיוון ש-N & B מפחית את הזמן בפיקסל למספר מיקרו-שניות, הוא יכול לעקוב אחר מהירות הקינטיקה המנותרת באזורי תאים גדולים, ולאפשר רכישת תמונה בסרגל זמן של שניות במיקרוסקופ סריקת רסטר (למשל, confocal וקד, 2-פוטון) ואלפיות השניה במיקרוסקופיה המבוססת על מצלמה (למשל, TIRFM).
מספר דוחות הפגינו את היכולת של N & B כדי לכמת את מספר יחידות המשנה באשכולות חלבונים על-ידי הדמיה של אזורי תאים מורחבים. אשכולות פעבלין-EGFP זוהו באתרי הדבקה ב-CHO-K1 תאים25, והצבירה התאיים של הפפטיד Httex1p הפתוגניים תוארה ב-COS-7 תאים26. N & B הוחל לאחר oligomerization מונחה ליגאן של הקולטן ErbB27, ואת ההשפעה של ליגFGF21 על קלותוב (klb) ו FGFR1c בתאי הלה28. השילוב של הדמיה TIRF ו-N & B ניתוח שימש כדי להראות כי דינמיות ב-2 היא בעיקר tetrameric לאורך כל קרום התא29. החלתי N & B לשני סריקת רסטר ותמונות tirf להוכיח dimerization מונחה ליגור של upar ו FGFR1 תא קולטנים קרום30,31.
שיטות מתאם פלואורסצנטית, כגון N & B, FCS ו-PCH, מבוססות על הרעיון שבנפח פתוח מספר הכיבוש של החלקיקים עוקב אחר התפלגות פואסון. כי רק את הפוטונים כי פולטות fluorophores ניתן להבחין, ערך ממוצע של עוצמת הזריחה נמדד לעומת הזמן בפיקסל של התמונה  ,, הוא התוצר של המספר הממוצע של fluorophores נפח התאורה, n, ואת ה בהירות מולקולרית, ε17:
,, הוא התוצר של המספר הממוצע של fluorophores נפח התאורה, n, ואת ה בהירות מולקולרית, ε17:

כאשר ε מבוטא כמספר הפוטונים הנפלטים ליחידת זמן (מקובל לשניה) לכל מולקולה כאשר המולקולה נמצאת במרכז עוצמת התאורה.
בהירות היא תכונה של כל fluorophore ברכישה נתונה להגדיר, בעוד האינטנסיביות הוא הסכום של כל התרומות מכל fluorophores. בתחרויות ביולוגיות, הבהירות יגדל עם הגידול של מספר fluorophores המתאחד יחד, נותן מידע על המצב oligomerization של חלבון fluoresc, מתויג. מגביר תנודות בפיקסל נתון נמדד מן השונות של האות הפלואורסצנטית, σ2:

כאשר המשמעות של ריבוע העוצמה  , והריבוע של העוצמה
, והריבוע של העוצמה  , מחושבים מערכי העוצמה האינדיווידואליים בכל פיקסל בכל מסגרת:
, מחושבים מערכי העוצמה האינדיווידואליים בכל פיקסל בכל מסגרת:
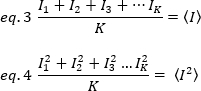
כאשר K הוא מספר המסגרות הכוללות בסידרת הזמן. ניסויים, יש צורך לחשב עבור סדרת התמונות כולה את השונות המתארת את פיזור ערכי העוצמה הבודדת בכל פיקסל של תמונה אחת סביב ערך העוצמה הממוצע. השונות כוללת את כל התנודות במקורות שונים. בקירוב ראשון, השונות הנובעת מהחלקיקים הניתנים לפיזור בעוצמת התאורה, σ20, יכולה להיות מופרדת מן השונות עקב רעש שוט הגלאי, σ2ד. שני השונויות הן עצמאיות; לפיכך, השונות הכוללת ניתנת על-ידי הסכום שלהם:

השונות, עקב תנודות מולקולריות של עוצמת האיתור והיציאה, תלויה בצורה לינארית בבהירות ובעוצמה המולקולרית:

מסידור מחודש של eq 6 לפי eq .1:

על פי המושג האופייני ספקטרוסקופיית מתאם פלואורסצנטית, משוואה 7 קובעת כי השונות בשל מספר התנודות תלויה בריבוע של בהירות החלקיקים.
לאחר מכן, השונות עקב תנודות גלאי היא פונקציה קווית של העוצמה שזוהתה, תחת ההנחה כי הגלאי מופעל מתחת למגבלת הרוויה שלה19:

במקרה של גלאים של ספירת פוטון a= 1 ו- c= 0, ולכן שונות הגלאי שווה לעוצמה הממוצעת:

כדי להחיל מושגים אלה על מדידות אמיתיות בתאים חיים, Gratton ועמיתים18 להגדיר את הבהירות לכאורה, B, עבור כל פיקסל כיחס של שונות על העוצמה הממוצעת:

B הוא הפרמטר הנמדד באמצעות הניסויים. בעבודה זו, סדרת הזמן תמונות של קולטני FGFR1 בקרום פלזמה של תאי הלה נלכדים על ידי מיקרוסקופ TIRF ואת הבהירות לכאורה הממוצעת, B, נקבעת על ידי ניתוח N & B. לאחר מכן, לאחר הוספת FGF2, סדרת הזמן ברציפות נלכדים לבצע את השינויים העצמית הרכבה של מולקולות הקולטן במשטח הקרום לאחר גירוי של הקולטן עם ligand קאנוני.
עם זאת, מאז גלאי המיקרוסקופ TIRF הוא מצלמה EMCCD, הביטוי לבהירות לכאורה צריך להיות שונה19:

כאשר היסט הוא היסט האינטנסיביות של האלקטרוניקה לאיתור שהוא מאפיין של הגדרות הגלאי. השונות והעוצמה הממוצעת עבור גלאי אנלוגי ניתנות בהתאמה על-ידי:
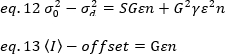
כאשר G הוא רווח אנלוגי ברמות דיגיטליות (DL/פוטונים), S, הרמות הדיגיטליות לכל פוטון19, ניתנת על ידי השיפוע של עוצמה לעומת מזימה שונות עבור מקור אור עם עוצמה מתמדת (אין תנודות בזמן). הפקטור הγ קשור לצורה של אמצעי האחסון לזיהוי פיקסלים. לדברי האסלר ואח ‘32, הפקטור γ שווה 0.3 עבור הדמיה tirf עבודה ברווח המקסימלי של מצלמת האיתור19. היסט, S ו-G פרמטרים הם מאפיינים של המצלמה ואת המיקרוסקופ. הבהירות לכאורה, B, מתקבלת על ידי סידור משנה של eq. 11 לפי eq .12 ו -13:

ניסויים, ε היא פונקציה מורכבת של עוצמת לייזר ויעילות הזיהוי של המערכת. עם זאת, מאחר ש-B/S תלוי באופן ליניארי ב ε, חשוב רק לקבוע את הערך היחסי של ε עבור מצב זיהוי נתון:

כאשר ε ‘ הוא פרופורציונלי לε. עדיין, כיול מבוצע באמצעות הפניה פנימית.
