La procedura per la correzione cromatica è un compromesso tra precisione e sforzo. Per risparmiare sforzi inutili, è meglio sapere quanta precisione è necessaria per il vostro studio. La massima precisione potrebbe non essere richiesta per l’imaging tradizionale a campo largo (dal vivo) e, pertanto, le immagini di riferimento luminose sono spesso sufficienti per correggere lo spostamento cromatico. Allo stesso modo, quando la condizione di imaging e l’ambiente sono costanti, l’uso ripetuto di una calibrazione biologica farà risparmiare tempo. D’altra parte, se si desidera una registrazione altamente accurata, sono necessarie immagini di riferimento trasversali o di calibrazione biologica di alta qualità. Per ottenere prestazioni ottimali, le immagini di riferimento devono essere ottenute con condizioni e tempi simili il più possibile le immagini di destinazione. Finché sia le immagini di riferimento che le immagini di destinazione sono ottenute dalla stessa microscopia, una maggiore risoluzione spaziale migliorerà la precisione di correzione. Se la deconvoluzione è disponibile sia per le immagini di riferimento che per le immagini di destinazione, l’implementazione di questa correzione prima della correzione può migliorare la precisione della correzione. Inoltre, per ottenere le migliori prestazioni, il teorema di campionamento per l’asse ottico (sezione) deve essere soddisfatto sia nel file di riferimento che in quello di destinazione per un’interpolazione precisa dei subpixel (passaggio del protocollo 2.1.3).
La mancata correzione del cambiamento cromatico porta a conclusioni errate. Inoltre, l’uso della calibrazione sbagliata può persino peggiorare i cambiamenti cromatici piuttosto che correggerli, e questo deve quindi essere evitato. Abbiamo riassunto le possibili cause degli errori e le loro soluzioni comuni nella tabella 2. Per esaminare la causa di un guasto, in primo luogo, è necessario controllare visivamente se lo spostamento cromatico nell’immagine di riferimento è corretto con precisione (passaggio del protocollo 3.12). La maggior parte degli errori sono dovuti alla qualità delle immagini di riferimento e sono facilmente risolvibili secondo le descrizioni nella tabella 2. Per quanto riguarda la qualità delle immagini di riferimento, è importante notare che la precisione dell’allineamento globale diminuisce se l’intero campo visivo non viene riempito con il campione (Figura 7, Tabella 2). Rispetto al buon esempio illustrato nella figura 7A, l’esempio non valido illustrato nella figura 7B contiene solo tre inviluppi nucleari nell’area superiore sinistra e Chromagnon non è riuscito ad allineare una parte di questa immagine. Questo perché il metodo di allineamento globale di Chromagnon divide il campo visivo in quattro regioni (Figura 7C) al fine di misurare le differenze di rotazione e ingrandimento con alta precisione3. Questo metodo, se correttamente utilizzato, è un ordine più preciso rispetto ad altri metodi lineari come la trasformazione polare del log e i metodi simplex3. Se una delle quattro regioni non è disponibile, Chromagnon passerà a metodi lineari meno efficaci. Pertanto, per ottenere prestazioni ottimali, gli esempi illustrati nella figura 7B e nella figura 7C sono indesiderati e le quattro aree devono essere riempite con oggetti. Gli utenti possono verificare se qualsiasi area quadratica del campo visivo non è disponibile per la misurazione esaminando il file di registro (“Chromagnon.log”; vedere il passaggio 3.10 del protocollo). Fortunatamente, questo problema può essere facilmente risolto calcolando la media di più immagini di calibrazione biologica o utilizzando l’allineamento locale per le immagini di riferimento crosstalk o bright-field (Tabella 2). Contrariamente al caso di mancata corretta delle immagini di riferimento, è più difficile identificare le immagini di destinazione. Poiché tali errori sorgono a causa di differenze nei formati di file, nelle condizioni di imaging, nei tempi di imaging, nei metodi di imaging/allineamento tra le immagini di riferimento e di destinazione (Tabella 2), gli utenti devono sempre prestare attenzione quando utilizzano immagini di riferimento ottenute in condizioni/intervalli diversi dalle immagini di destinazione. Alcune immagini di esempio sono disponibili per il test (https://github.com/macronucleus/Chromagnon) per ottenere un’idea concreta delle immagini di esempio buone e cattive.
| Problema | Causa | Soluzione |
| Impossibile correggere l’immagine di riferimento | Basso contrasto | Se possibile, acquisire un’immagine a contrasto più elevato. Se viene utilizzata un’immagine di riferimento di campo luminoso, riacquisire l’immagine in una soluzione basata sull’acqua per ottenere un contrasto maggiore della cella. In alternativa, provare ad applicare la riduzione del rumore computazionale (ad esempio il filtraggio gaussiano). Disattivare l’allineamento locale, che è più sensibile al disturbo. |
| Contaminazione di immagini non correlate | Se possibile, rimuovere l’origine delle immagini non correlate nell’esempio. Per le immagini di riferimento tra gambo, controllare gli spettri di eccitazione dei coloranti utilizzati per le immagini di destinazione. Se i coloranti sono eccitati durante l’acquisizione di un’immagine crosstalk (ad esempio Alexa Fluor 568 o 594), prendere in considerazione altri coloranti (ad esempio Alexa Fluor 555). Se le polveri sul chip della fotocamera creano un’ovvia differenza di canale, pulire il chip della fotocamera o utilizzare un metodo di applicazione a campo piatto computazionale. | |
| Un punto estremamente luminoso fatto da un raggio cosmico | Se possibile, acquisire nuovamente l’immagine. In alternativa, provate ad applicare la riduzione del rumore computazionale (ad es. filtro mediano o gaussiano). | |
| Artefatti di deconvoluzione (segnali artificiali ai bordi assiale e laterale) | Tagliare i pixel del bordo o le sezioni , dopo la deconvoluzione. Se un lato è tagliato, anche l’altro lato deve essere tagliato per mantenere il centro dell’immagine. | |
| La dimensione del passo di z troppo scarsa | Per soddisfare il criterio nyquist, come indicato nel protocollo 2.1.3, è necessario acquisire uno stack di tipo z. | |
| Aberrazione ottica | L’aberrazione sferica è la principale aberrazione causata dagli utenti. Scegliere l’obiettivo giusto per il campione e utilizzare uno spessore coverslip di 170 m. Se l’obiettivo è dotato di un anello di correzione, regolarlo per trovare la posizione in cui si ottiene il numero di fluorescenza più alto dalla messa a fuoco. Nel caso di un obiettivo di immersione dell’olio senza un anello di correzione, regolare l’indice di rifrazione dell’olio di immersione che aumenta il conteggio della fluorescenza al fuoco. | |
| Il campo visivo non è riempito (Fig. 7) | Nel caso delle immagini di riferimento della calibrazione biologica, mediamente molte immagini. Nel caso di immagini a gambo incrociato o a campo luminoso, utilizzare l’allineamento locale. | |
| Un bug software non identificato | Segnalare il problema tramite GitHub(https://github.com/macronucleus/Chromagnon/issues) | |
| Impossibile correggere l’immagine di destinazione | I metadati del file di immagine vengono persi | Utilizzare il formato di file dell’ambito originale che contiene metadati completi ed evitare la conversione in un file tiff a più pagine prima dell’elaborazione. Utilizzare lo stesso ordine dei canali come scritto nel protocollo 3.3. |
| Metodi di allineamento errati per la microscopia data | Non applicare il metodo di allineamento locale durante la misurazione da immagini di riferimento di calibrazione biologica alle immagini di destinazione. Non utilizzare immagini di riferimento a gambo incrociato diverse dalla microscopia a campo largo. | |
| Differenze nelle condizioni di imaging | Mantenere costanti le condizioni di imaging tra le immagini di riferimento e di destinazione come scritte nel protocollo 2.3.3. | |
| Differenze nel campione (compreso coverslip) | Utilizzare sempre lo stesso supporto di montaggio, coverslip (ad es. n. 1.5H) e una profondità di messa a fuoco simile. | |
| Deriva del microscopio dall’ultima | Effettuare una calibrazione tutte le volte ogni due settimane. Mantenere la temperatura costante e utilizzare una tabella mobile per evitare la deriva hardware del microscopio. |
Tabella 2: Risoluzione dei problemi relativi alla correzione cromatica.
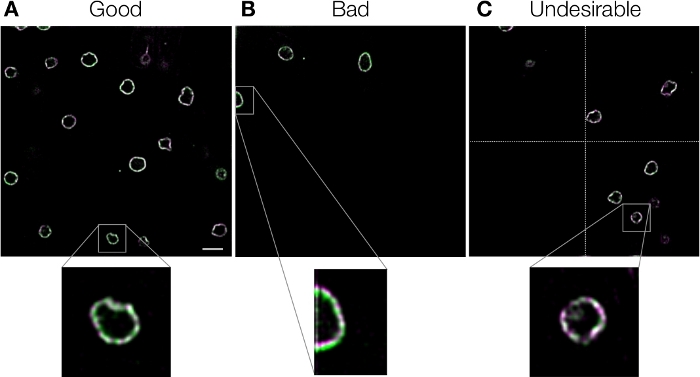
Figura 7: Esempi di immagini di riferimento. Busta nucleare in celle di lievito di fissione etichettate con GFP e mCherry. Le immagini sono state acquisite con la microscopia a campo largo convenzionale. I cambiamenti cromatici sono stati corretti utilizzando Chromagnon senza allineamento locale utilizzando le immagini stesse come immagini di riferimento. Le immagini sono state poi deconvolved per mostrare i dettagli. (A) Un buon esempio con molti oggetti nel campo visivo. (B) Un cattivo esempio con gli oggetti solo nell’angolo in alto a sinistra. Il disallineamento è evidente in una determinata regione dell’immagine. (C) Un esempio indesiderato in cui una delle quadrisezioni (separate da linee trasversali tratteggiate) è vuota. La barra della scala nel pannello A indica 5 m per la vista a campo intero e 1,25 m per la vista ingrandita ed è applicabile a tutti i pannelli. Fare clic qui per visualizzare una versione più grande di questa figura.
In questo protocollo sono stati descritti tre diversi tipi di riferimento(Tabella 1). Tra questi, le immagini di riferimento tra le gambo incrociate e le immagini di riferimento di calibrazione biologica necessitano di un’ulteriore attenta discussione. Per le immagini di riferimento ai concrologi, i campioni macchiati con DAPI o Hoechst 33342 e montati in glicerolo o supporti di montaggio commerciali possono essere utilizzati in modo efficiente per allineare i canali blu, verde e rosso. Allo stesso modo, Alexa Fluor 488 può essere utilizzato per allineare i canali verde e rosso. Tuttavia, ottenere la fluorescenza trasversale è spesso difficile dal momento che molti coloranti blu tranne DAPI e Hoechst sono dimmer e decadimento più velocemente rispetto alla maggior parte coloranti verdi e rossi. Inoltre, gli spettri di emissione dei coloranti moderni sono più stretti, il che rende difficile l’allineamento di più di tre canali con questo metodo. Attenzione dovrebbe essere prestata anche ad alcuni coloranti rossi comuni (ad esempio, Alexa Flour 568 e 594, ma non Alexa Fluor 555) che possono essere eccitati dalla luce viola, che impediscono di ottenere immagini di crosstalk ad alto contrasto da coloranti blu. Un altro inconveniente è che questo metodo non può misurare l’aberrazione cromatica dei percorsi di luce di eccitazione in eccitazione multicolore, perché solo una singola lunghezza d’onda di eccitazione viene utilizzata per l’eccitazione (Tabella 1). Poiché la microscopia più avanzata utilizza un’ottica di illuminazione alterata, l’applicazione di questo metodo è limitata. Tuttavia, la maggiore precisione di correzione è sufficientemente vantaggiosa per essere descritta in questo protocollo. In generale, un’immagine crosstalk deve essere scattata dopo un’immagine di destinazione per evitare effetti sbiancanti o fototossici. Per SMLM osservato con la modalità wide-field, un’immagine di riferimento deve essere acquisita prima di acquisire un’immagine di destinazione come coloranti a fluorescenza può essere sbiancata durante l’imaging.
Le immagini di riferimento della taratura biologica consentono agli utenti di allineare facilmente qualsiasi numero desiderato di canali a costo di una preparazione aggiuntiva del campione. Un altro vantaggio delle immagini di riferimento della taratura biologica è la disponibilità di “media” più riferimenti che aiuta a riempire tutti i campi visivi. Questo metodo può soffrire di differenze nelle condizioni di imaging se il campione di calibrazione viene preparato su un vetrino diverso. La maggior parte di questo problema può essere risolta se sia le destinazioni che i riferimenti vengono preparati sulla stessa diapositiva utilizzando le coperture a camere commerciali (Tabella 1) e altre condizioni di imaging sono mantenute costanti come nel passaggio di protocollo 2.3.3. In questo caso, ci si può aspettare una precisione di correzione simile a quella delle immagini di riferimento del gambo incrociato3. Il protocollo per l’utilizzo della falloidina come mostrato qui è uno dei modi più semplici per macchiare una singola struttura cellulare con più colori. Ci sono numerosi scenari possibili per preparare campioni di calibrazione biologica. Per l’immunostaining, un campione può essere etichettato con un singolo anticorpo primario seguito da colorazione con anticorpi secondari di più colori. In questo modo, una singola struttura di destinazione può essere etichettata con più colori. In alternativa, 5-ethynyl-2′-deoxyuridina, rilevata da etichette chimiche “click” appena sintetizzate DNA in più colori ad alta densità, come descritto nel dettaglio precedentemente8. Per le cellule vive, è utile preparare un ceppo transgenico che ospita due copie di un gene che vengono fuse in GFP o mCherry per etichettare la stessa struttura con due colori. Se il numero di copie del gene è critico come spesso osservato per le proteine della membrana, una singola copia del gene può essere tandemly fusa a GFP e mCherry (Figura 7). Le proteine fluorescenti fotoconvertibili, come mEOS218, possono essere utilizzate anche illuminando un livello moderato di luce viola per ottenere entrambe le specie proteiche con o senza fotoconversione. In basse condizioni di ossigeno, GFP può anche essere utilizzato come proteina fotoconvertibile dal verde al rosso19,20. La scelta del campione di calibrazione corretto renderà quindi l’esperimento più robusto.
