غالبًا ما تتكون تحقيقات التمثيل الغذائي الثانوي من فحص المستخلصات الخام لأنشطة بيولوجية محددة يتبعها تنقية وتحديد وتوصيف المكونات التي تنتمي إلى كسور نشطة. وقد أثبتت هذه العملية كفاءتها، مما يعزز عزل العديد من الكيانات الكيميائية. ومع ذلك ، في الوقت الحاضر يعتبر هذا غير ممكن ، ويرجع ذلك أساسا إلى ارتفاع معدلات إعادة اكتشاف. وبما أن صناعة الأدوية أحدثت ثورة دون معرفة أدوار ووظائف الأيض المتخصصة، فقد تم تحديدها في ظروف مختبرية لا تمثل الطبيعة بدقة1. اليوم ، هناك فهم أفضل لتأثيرات الإشارات الطبيعية ، والإفراز ، ووجود معظم الأهداف بتركيزات منخفضة بشكل لا يمكن اكتشافه. بالإضافة إلى ذلك، سيساعد تنظيم العملية المجتمع الأكاديمي وصناعة الأدوية على الاستفادة من هذه المعرفة. كما سيفيد البحوث التي تنطوي على العزل المباشر للمستقلبات المتعلقة بمجموعات الجينات الاصطناعية البيولوجية الصامتة (BGCs)2.
وفي هذا السياق، جدد التقدم في التسلسل الجينومي الاهتمام بفرز نواتج الأيض الدقيقة. وذلك لأن تحليل المعلومات الجينومية لمجموعات التركيب الحيوي المكشوف ة يمكن أن يكشف عن الجينات ترميز المركبات الجديدة لا لوحظ أو تنتج في ظل ظروف المختبر. تتوفر العديد من مشاريع الجينوم الميكروبية الكاملة أو المسودات اليوم ، ويتزايد العدد كل عام ، مما يوفر آفاقًا هائلة للكشف عن جزيئات جديدة نشطة بيولوجيًا من خلال تعدين الجينوم3،4.
أطلس مجموعات الجينات التركيبية الحيوية هو أكبر مجموعة حالية من مجموعات الجينات الملغومة تلقائيًا كمكون من منصة الجينوم الميكروبي المتكامل لمعهد الجينوم المشترك (JGI IMG-ABC)2. وفي الآونة الأخيرة، عززت مبادرة توحيد مجموعات الجينات الاصطناعية الأحيائية الحد الأدنى من المعلومات (MIBiG) الشرح اليدوي لـ BGCs، مما يوفر مجموعة بيانات مرجعية منسقة للغاية5. في الوقت الحاضر ، تتوفر الكثير من الأدوات لتمكين التعدين الحسابي للبيانات الوراثية واتصالها بالأيض الثانوي المعروف. كما تم وضع استراتيجيات مختلفة للوصول إلى منتجات طبيعية جديدة نشطة بيولوجياً (أي التعبير غير المتجانس، وحذف الجينات المستهدفة، وإعادة التشكيل المختبري، والتسلسل الجينومي، والفحص الموجه بالنظائر [نهج جنوميسوتوبي]، والتلاعب بالمنظمين المحليين والعالميين، والتعدين القائم على الأهداف المقاومة، والتعدين المستقل للثقافة، ومؤخراً،نهجMS/code الموجهة2،6،,77،,8،9،, 10،11،12،13،14،15).
يتطلب تعدين الجينوم كاستراتيجية فريدة بذل جهود لتعليقات توضيحية لمجموعة واحدة أو صغيرة من الجزيئات. وبالتالي، لا تزال هناك ثغرات في العملية التي تعطى فيها الأولوية للمركبات الجديدة من أجل عزلها وتوضيح هيكلها. ومن حيث المبدأ، لا تستهدف هذه النهج سوى مسار واحد للتركيب الحيوي لكل تجربة، مما يؤدي إلى بطء معدل الاكتشاف. في هذا المعنى ، فإن استخدام جنرال موتورز جنبا إلى جنب مع نهج الشبكات الجزيئية يمثل تقدما هاما لبحوث المنتجات الطبيعية14،15.
إن تعدد الاستخدامات والدقة والحساسية العالية لقياس الطيف اللوني الكتلي السائل (LC-MS) يجعلها طريقة جيدة لتحديد المركب. حاليا، وقد استثمرت العديد من المنصات الخوارزميات وأجنحة البرمجيات لmetabolomics غير المستهدفة16،,17،,18،,19، 20,20. يتضمن جوهر هذه البرامج اكتشاف الميزات (ذروة الانتقاء)21 ومحاذاة الذروة ، مما يسمح بمطابقة الميزات المتطابقة عبر مجموعة من العينات والبحث عن الأنماط. الخوارزميات المستندة إلى نمط MS22،23 مقارنة أنماط التجزئة المميزة وتطابق أوجه التشابه MS2 التي تولد العائلات الجزيئية التي تشترك في الميزات الهيكلية. ويمكن بعد ذلك تسليط الضوء على هذه الميزات وتتجمع، ومنح القدرة على اكتشاف بسرعة جزيئات2معروفة وغير معروفة من استخراج البيولوجية المعقدة من قبل MS جنبا إلى جنب2،24،,25. ولذلك، فإن التصلب المتعدد المترادف هو طريقة متعددة الاستخدامات للحصول على معلومات هيكلية عن العديد من الأنماط الكيميائية الواردة في كمية كبيرة من البيانات في وقت واحد.
تستخدم خوارزمية الشبكات الجزيئية الاجتماعية للمنتجات الطبيعية العالمية (GNPS)26 كثافة الأيونات المجزأة العادية لبناء ناقلات متعددة الأبعاد ، حيث تتم مقارنة أوجه التشابه باستخدام وظيفة جيب الجيب. يتم رسم العلاقة بين الأيونات الأصلية المختلفة في تمثيل مخطط، حيث يتم تصور كل تجزئة كعقدة (دوائر)، ويتم تعريف الصلة بين كل عقدة بواسطة حافة (خطوط). يتم تعريف التصور العالمي للجزيئات من مصدر واحد كشبكة جزيئية. الجزيئات المتباينة هيكليا التي تتفتت بشكل فريد ستشكل مجموعة خاصة بهم أو كوكبة، في حين أن الجزيئات ذات الصلة تتجمع معا. يسمح الأنماط الكيميائية العنقودية باتصال افتراضي من الميزات الهيكلية المماثلة لأصولها التركيبية الحيوية.
الجمع بين كل من النمط الكيميائي إلى النمط الجيني ونهج النمط الجيني إلى النمط الكيميائي هو قوي عند إنشاء روابط المعلوماتية الحيوية بين BGCs ومنتجاتها جزيء صغير27. ولذلك، فإن تعدين الجينوم الموجه بالتصلب المتعدد هو طريقة سريعة واستراتيجية منخفضة الاستهلاك للمواد، ويساعد على سد الأيون اتّصال الوالدين والمسارات الاصطناعية الحيوية التي كشفت عنها WGS لسلالة واحدة أو أكثر في ظل ظروف استقلابية وبيئية متنوعة.
سير العمل من هذا البروتوكول(الشكل 1)يتكون من تغذية البيانات WGS في منصة الشرح الكتلة الجينية الاصطناعية مثل antiSMASH28،29،30. فهو يساعد على تقدير مجموعة متنوعة من المركبات وفئة من المركبات المشفرة من قبل الجينوم. يجب اعتماد استراتيجية لاستهداف مجموعة الجينات الاصطناعية الحيوية ترميز كيان كيميائي ذات فائدة، ومقتطفات الثقافة من سلالة نوع البرية و / أو سلالة غير متجانسة تحتوي على BGC يمكن تحليلها لتوليد الأيونات المجمعة على أساس أوجه التشابه باستخدام GNPS26،31. وبالتالي، فمن الممكن تحديد جزيئات جديدة ترتبط بـ BGC المستهدفة وغير متوفرة في قاعدة البيانات (نظائرها غير معروفة بشكل رئيسي، وتنتج في بعض الأحيان في تاتر منخفضة). ومن المهم النظر في أن المستخدمين يمكن أن تسهم في هذه المنصات وأن توافر المعلوماتية الحيوية وبيانات MS / MS يتزايد بسرعة ، مما يؤدي إلى تطوير مستمر ورفع مستوى الأدوات الحسابية الفعالة والخوارزميات لتوجيه اتصالات فعالة من مقتطفات معقدة مع الجزيئات.
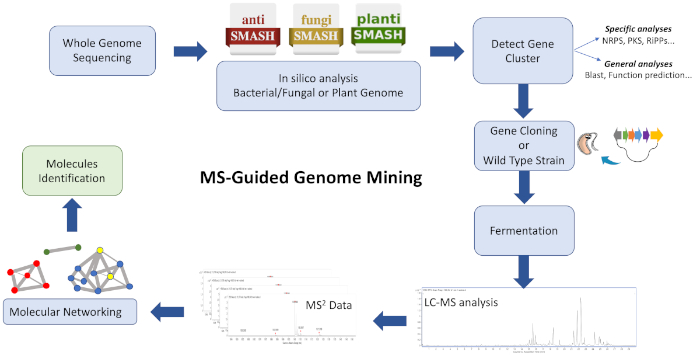
الشكل 1: نظرة عامة على سير العمل بأكمله. يُظهر هذا الشكل توضيحًا لخطوات المعلوماتية الحيوية والاستنساخ والشبكات الجزيئية التي تنطوي عليها نهج تعدين الجينوم الموجه بالتصلب المتعدد الموصوف لتحديد الأيض الجديدة. يرجى الضغط هنا لعرض نسخة أكبر من هذا الرقم.
يصف هذا البروتوكول سير العمل السريع والفعال للجمع بين تعدين الجينوم والشبكات الجزيئية كنقطة انطلاق لخط أنابيب اكتشاف المنتج الطبيعي. على الرغم من أن العديد من التطبيقات قادرة على تصور تكوين وعلاقة الجزيئات التي يمكن اكتشافها بالتصلب المتعدد في شبكة واحدة ، يتم اعتماد العديد منها هنا لتصور جزيئات مجمعة متشابهة هيكلياً. باستخدام هذه الاستراتيجية، يتم تحديد منتجات سيكلوبسيبتيد الرواية التي لوحظت في المستخلصات الأيضية من Streptomyces sp. CBMAI 2042 بنجاح. تسترشد التعدين الجينوم، يتم التعرف على كامل ترميز الكتلة الجينية الاصطناعية الحيوية لفالينوميسين واستنساخها في سلالة المنتج Streptomyces coelicolor M1146. وأخيراً، وبعد إقامة الشبكات الجزيئية القائمة على نمط MS، ترتبط الجزيئات التي تم اكتشافها بواسطة التصلب المتعدد بـ BGCs المسؤولة عن تكوينها الحيوي32.
