Impulsada por la creciente aplicación de técnicas de secuenciación de próxima generación, se podría aumentar la resolución espacial en la biología del desarrollo. Los estudios contemporáneos tienen como objetivo disuajar tejidos hasta tipos de células especializadas, si no de un solo nivel de células1,,2,3,4. Para ello, en los últimos cincuenta años se ha ideado una plétora de métodos diferentes (véase la Figura 1A)5,6,7,8,9,10,11,12,13,14,15.
Muchas herramientas en la ciencia vegetal han sido adaptaciones de técnicas que fueron pioneras en la investigación animal. Este no es el caso del método que estamos introduciendo en detalle aquí. En 2005, equipado con una sólida experiencia en traducción de proteínas, el Bailey-Serres Lab se propuso diseñar proteínas ribosomales para la posterior purificación de afinidad16. Por lo tanto, podrían evitar el perfilado de polisomas que consume mucho tiempo y requiere mucha mano de obra, que se basa en la ultracentrifugación con un gradiente de sacarosa y se utilizó para evaluar la traducción de ribosomas desde la década de 196017,18. Desde entonces, el método se ha denominado purificación de afinidad ribosoma traslacional (TRAP)16. Después de estudios de traducción exitosos en plantas, Heiman et al. adaptó TRAP para animales19 y otros ampliaron su aplicación a la levadura20, Drosophila21, Xenopus22 y zebrafish23,,24.
Aunque la modificación genética del sistema modelo es un requisito previo para trap, que limita su aplicación a especies susceptibles a la transformación genética, se puede aprovechar simultáneamente esta objeción a los subconjuntos objetivo de células que son de especial interés y, por lo tanto, extremadamente difíciles de aislar del tejido/órgano intacto25 (por ejemplo, células dendríticas altamente ramificadas en un cerebro de ratón o hifas fúngicas en el tejido vegetal infectado). En las plantas, todas las células se mantienen en su lugar a través de paredes celulares que forman la base del esqueleto hidrostático26. Para liberar una célula vegetal de esta matriz, los científicos han cortado físicamente la célula de su tejido circundante a través de la microdisección de captura láser (LCM)27 o han realizado digestión enzimática de las paredes celulares28. Entre estas últimas células, las llamadas protoplastias, la población de interés está etiquetada fluorescentemente y se puede separar mediante la clasificación celular activada por fluorescencia (FACS)7. LCM generalmente requiere que una muestra sea fija e incrustada en la cera, lo que en última instancia deteriora la calidad de su ARN29. Los métodos basados en FACS producen ARN de alta calidad, pero el proceso de protoplasting en sí mismo introduce diferencias en la expresión génica30 y los tejidos con paredes celulares secundarias modificadas y gruesas son notoriamente difíciles de tratar. Además, se supone que muchos procesos de desarrollo en las plantas dependen de señales transmitidas mecánicamente y, por lo tanto, la integridad de la pared celular es de suma importancia31. Dos métodos, que utilizan un acceso directo para eludir el aislamiento celular operando en el nivel de nucleii, son la clasificación nuclear activada por fluorescencia (FANS) y el aislamiento de núcleos etiquetados en tipos de células específicos (INTACT). Al igual que en TRAP, utilizan promotores específicos del tipo de celda para marcar los núcleos, que posteriormente se enriquecen a través de la clasificación o el tirón hacia abajo, respectivamente8,15. Un desafío importante para todos estos enfoques es obtener suficiente material de ARN de subconjuntos de células en un tejido. Como TRAP captura solo una fracción de los ARN celulares, la recolección de muestras es un cuello de botella considerable. Por lo tanto, se necesitan protocolos de preparación de bibliotecas especialmente sensibles para producir datos de alta calidad a partir de cantidades de entrada bajas.
Desde su creación, TRAP se ha utilizado en combinación con microarrays de ADN o, como los costos de secuenciación disminuyeron significativamente en los últimos años, ARN-seq10,32,33. Una multitud de preguntas de investigación ya se ha aclarado como se revisó en Sablok et al.34. Estamos convencidos de que seguirán más informes en los próximos años, ya que la técnica es muy versátil a la hora de combinar diferentes promotores para apuntar a tipos de células específicas. Eventualmente, esto se hará incluso de una manera inducible, y puede combinarse con el sondeo de la reacción de la planta a muchos factores de estrés biótico y abiótico. Además, cuando no se dispone de líneas transgénicas estables, los sistemas de expresión de raíces peludas también se han utilizado con éxito para realizar TRAP en tomate y medicago35,,36.
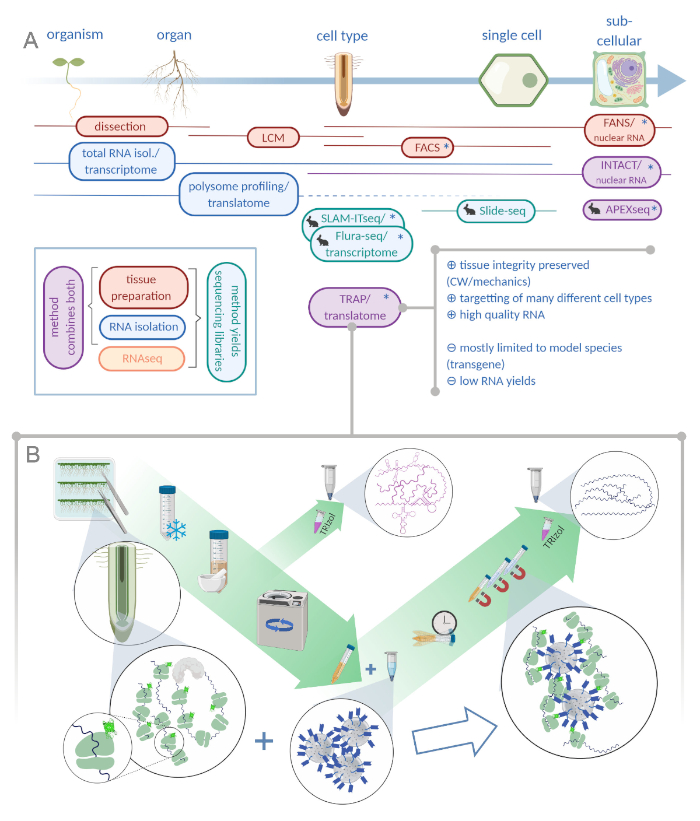
Figura 1: La traducción de la purificación de afinidad ribosoma (TRAP) complementa la cartera de análisis “omics”. A. El aumento de los niveles de precisión analítica, hasta una sola célula o incluso la resolución subcelular se puede lograr mediante una plétora de métodos o combinaciones de los mismos. El esquema ofrece una visión general de las herramientas disponibles actualmente en el campo vegetal y animal. La recolección de tejidos a resolución celular se puede lograr mediante protocolos como LCM o FACS, que luego se acoplan al transcriptoma estándar o al análisis de perfiles/traducción de polisoma. TRAP e INTACT integran la captura de tejido y el aislamiento de ARN, ya que se basan en el etiquetado de epítopos. Sin embargo, INTACT sólo toma muestras de núcleos celulares y constituye, por lo tanto, un caso especial de análisis de transcriptoma. Un pequeño icono de conejo marca los métodos recientemente desarrollados en el campo animal: Mientras que SLAM-ITseq y Flura-seq se basan en el objetivo metabólico de ARN nacientes con bases de uracilo modificadas en células que expresan la enzima permisiva, Slide-seq hace uso de un portaobjetos de vidrio recubierto con códigos de barras de ADN que proporcionan información posicional en el rango celular. Se sigue un enfoque de etiquetado de proximidad en APEX-seq para muestrear ARN en compartimentos subcelulares específicos. En particular, el aumento de la resolución a menudo requiere la generación de material transgénico (asteriscos) y, por lo tanto, estos métodos se utilizan predominantemente para las especies modelo. TRAP es especialmente adecuado para estudios de ciencias vegetales que involucran pared celular (CW) o señalización mecánica, así como especies celulares que son difíciles de liberar de su matriz CW. B. Pasos detallados de laboratorio húmedo del procedimiento TRAP: Las plántulas que expresan proteína ribosomal etiquetada con GFP en distintos tipos de células (por ejemplo, endodermis de raíz) se cultivan en platos De Petri durante siete días y el material radicular cosechado por congelación rápida. Se recoge una muestra de control total de ARN del extracto crudo homogeneizado antes de peletizar los escombros a través de la centrifugación. Las perlas magnéticas anti-GFP se añaden al extracto transparente para realizar la inmunoprecipitación. Después de la incubación y tres pasos de lavado, el ARN asociado al polisoma (ARN TRAP/polisoma) se obtiene directamente a través de la extracción de fenol-cloroformo. LCM: microdisección de captura láser, FACS/FANS: clasificación celular/nuclear activada por fluorescencia, APEX-seq: método basado en ascorbación peroxidasa diseñada, INTACT: aislamiento de núcleos etiquetados en tipos de células específicas, alquilación vinculada a SLAM-ITseq: thiol(SH) para la secuenciación metabólica de ARN en tejido, Flura-seq: secuenciación de ARN con etiqueta de fluorouracilo (Creado con Biorender.com) Haga clic aquí para ver una versión más grande de esta figura.
El objetivo de este artículo es proporcionar una descripción detallada del método TRAP, resaltar los pasos críticos y proporcionar orientación para un posible método de preparación de biblioteca.
Un experimento trap genérico consistirá esencialmente en los siguientes pasos (véase también la figura 1B):(1) Preparación de material vegetal, incluida la clonación de la construcción de etiquetado de ribosoma, producción y selección de líneas transgénicas, cultivo y abultamiento de semillas, esterilización y chapado, y aplicación/tratamiento de tensión (opcional) y recolección de tejidos; 2) inmunopurificación, incluida la homogeneización tisular y la limpieza del extracto crudo, el lavado de cuentas y la inmunopurificación, y las etapas de lavado; 3) extracción de ARN y evaluación de la calidad; y (4) la preparación de la biblioteca.
La raíz de Arabidopsis ha sido un sistema modelo para estudiar el desarrollo de la planta desde su introducción como planta modelo37,,38. Aquí, la aplicación de TRAP se muestra en el contexto del desarrollo de la raíz lateral de la planta. En las plantas, la acumulación de todo el sistema radicular se basa en la ejecución de este programa y por lo tanto es muy importante para la supervivencia del organismo39. En Arabidopsis,las raíces laterales se originan a partir de tejido periciclo que reside junto a vasos xilemas y, por lo tanto, se denomina periciclo de polo xilem (XPP; véase la figura 2C)40. Algunas células XPP, que se encuentran en lo profundo de la raíz, adquieren una identidad celular fundadora y, tras un desencadenante hormonal local, comienzan a proliferar por hinchazón y división anticlinalmente41. Sin embargo, debido a la presencia de una matriz de pared celular rígida, este proceso ejerce tensión mecánica en los tejidos circundantes. En particular, la endodermis superpuesta se ve afectada, ya que se encuentra en el camino del eje de crecimiento de la raíz lateral42,,43,44. De hecho, el primordium recién formado tendrá que crecer a través de la célula de endodermis superpuesta(Figura 2C2),mientras que las células de corteza y epidermis simplemente se apartan para que el primordium finalmente surja45,46. El trabajo reciente en nuestro laboratorio ha demostrado que la endodermis está contribuyendo activamente a acomodar la proliferación en el periciclo. El bloqueo dirigido de la señalización hormonal endodérmica es suficiente para inhibir incluso la primera división en las células XPP47. Por lo tanto, la comunicación periciclo-endodermis constituye un punto de control muy temprano para el desarrollo de la raíz lateral en Arabidopsis. Sin embargo, no se sabe cómo se realiza esta charla cruzada. Para desentrañar este misterio, elegimos el enfoque TRAP-seq para apuntar a las células XPP y endodeérmicas. Para enriquecer las células en el programa de raíz lateral, imitamos el desencadenante hormonal aplicando exógenamente un análogo de auxina (ácido 1-naftalencia, NAA)48,que al mismo tiempo permitió resolver temporalmente la fase inicial de la formación de la raíz lateral.
