Gedreven door de toenemende toepassing van next-generation sequencing technieken, ruimtelijke resolutie in de ontwikkelingsbiologie zou kunnen worden uitgebreid. Hedendaagse studies zijn gericht op het ontleden van weefsels tot gespecialiseerde celtypes, zo niet eencellige niveau1,2,3,4. Daartoe is in de afgelopen vijftig jaar een overvloed aan verschillende methoden bedacht (zie figuur 1A)5,6,7,8,9,11,11,12,13,14,15.
Veel instrumenten in de plantenwetenschap zijn aanpassingen van technieken die werden pionier in dierlijk onderzoek. Dit is niet het geval voor de methode die we hier in detail introduceren. In 2005, uitgerust met een sterke achtergrond in eiwitvertaling, ging het Bailey-Serres Lab op zoek naar ribosomale eiwitten voor latere affiniteitszuivering16. Zo konden ze tijdrovende en arbeidsintensieve polysome profilering vermijden, die is gebaseerd op ultracentrifugatie met een sacharosegradiënt en werd gebruikt om het vertalen van ribosomen te beoordelen sinds de jaren 196017,18. De methode is sindsdien aangeduid als translationele ribosoom affiniteit zuivering (TRAP)16. Na succesvolle translationoomstudies in planten, hebben Heiman et al. TRAP voor dieren19 en anderen de toepassing ervan uitgebreid tot gist20, Drosophila21, Xenopus22 en zebrafish23,24.
Hoewel genetische modificatie van het modelsysteem een voorwaarde is voor TRAP, dat de toepassing ervan beperkt tot soorten die vatbaar zijn voor genetische transformatie, kan men dit bezwaar tegelijkertijd benutten om subgroepen van cellen aan te pakken die van bijzonder belang zijn en anderszins uiterst moeilijk te isoleren van het intacte weefsel/orgaan25 (bijvoorbeeld sterk vertakte dendritische cellen in een muishersenof schimmelhyphae in besmet plantaardig weefsel). In planten worden alle cellen op hun plaats gehouden via celwanden die de basis vormen van het hydrostatische skelet26. Om een plantencel uit deze matrix te bevrijden, hebben wetenschappers de cel fysiek uit zijn omringende weefsel gesneden door middel van lasercapture microdissectie (LCM)27 of enzymatische spijsvertering van de celwandenuitgevoerd 28. Onder de laatste cellen, de zogenaamde protoplasten, is de populatie van belang fluorescerend gelabeld en kan worden gescheiden via fluorescentie-geactiveerde celsortering (FACS)7. LCM vereist meestal een monster te worden vastgesteld en ingebed in was, die uiteindelijk verslechtert de kwaliteit van haar RNA29. FACS-gebaseerde methoden leveren rna van hoge kwaliteit op, maar het proces van protoplasting zelf introduceert verschillen in genexpressie30 en weefsels met gemodificeerde en dikke secundaire celwanden zijn notoir moeilijk te behandelen. Bovendien wordt aangenomen dat veel ontwikkelingsprocessen in installaties afhankelijk zijn van mechanisch overgebrachte signalen en daarom is de integriteit van de celwand van het grootste belang31. Twee methoden, die een snelkoppeling gebruiken om celisolatie te omzeilen door te werken op het niveau van nucleii, zijn fluorescentie-geactiveerde nucleaire sortering (FANS) en isolatie van kernen die zijn getagd in specifieke celtypen (INTACT). Net als in TRAP gebruiken ze celtypespecifieke promotors om kernen te markeren, die vervolgens verrijkt worden via sorteren of naar beneden trekken, respectievelijk8,15. Een grote uitdaging voor al deze benaderingen is om voldoende RNA-materiaal te krijgen uit subsets van cellen in een weefsel. Aangezien TRAP slechts een fractie van de cellulaire RNA’s vangt, is monsterverzameling een aanzienlijk knelpunt. Daarom zijn er vooral gevoelige protocollen voor de voorbereiding van bibliotheken nodig om gegevens van hoge kwaliteit te produceren van lage invoerhoeveelheden.
Sinds de oprichting is TRAP ofwel gebruikt in combinatie met DNA-microarrays of, aangezien de sequencingkosten de afgelopen jaren aanzienlijk daalden, RNA-seq10,32,33. Een veelheid van onderzoeksvragen is al opgehelderd zoals beoordeeld in Sablok et al.34. We zijn ervan overtuigd dat er de komende jaren meer rapporten zullen volgen, omdat de techniek zeer veelzijdig is bij het combineren van verschillende promotors om specifieke celtypen te targeten. Uiteindelijk zal dit zelfs op een ondoordringbare manier gebeuren, en kan worden gecombineerd met het indringende van de reactie van de plant op vele biotische en abiotische stressfactoren. Bovendien, waar stabiele transgene lijnen niet beschikbaar zijn, zijn harige wortelexpressiesystemen ook met succes gebruikt om TRAP uit te voeren in tomaat en medicago35,36.
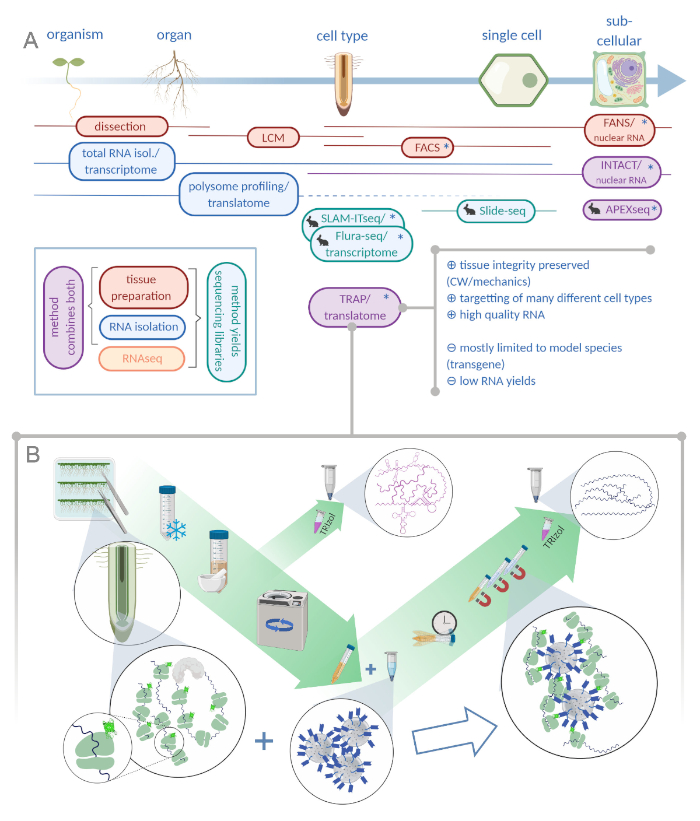
Figuur 1: Het vertalen van ribosoomaffiniteitszuivering (TRAP) vormt een aanvulling op het “omics” analyseportfolio. A. Het verhogen van de mate van analytische precisie, tot eencellige of zelfs subcellulaire resolutie kan worden bereikt door een overvloed aan methoden of combinaties daarvan. De regeling geeft een overzicht van de momenteel beschikbare gereedschappen op het gebied van planten en dieren. Weefselverzameling bij cellulaire resolutie kan worden bereikt door protocollen zoals LCM of FACS, die vervolgens worden gekoppeld aan standaard transcriptome of polysome profiling /translatome analyse. TRAP en INTACT integreren zowel weefselopname als RNA-isolatie omdat ze gebaseerd zijn op epitoop-tagging. INTACT monstert echter alleen celkernen en vormt daarom een speciaal geval van transcriptoomanalyse. Een klein konijn pictogram markeert nieuw ontwikkelde methoden in het gebied van dieren: Terwijl SLAM-ITseq en Flura-seq vertrouwen op metabole targetting van ontluikende RNAs met gemodificeerde uracil bases in cellen uitdrukken van de tolerante enzym, Slide-seq maakt gebruik van een gecoate glazen dia met DNA barcodes die positionele informatie in het cellulaire bereik. Een proximity labeling benadering wordt gevolgd in APEX-seq om RNA’s te proeven in specifieke subcellulaire compartimenten. Met name een verhoogde resolutie vereist vaak de productie van transgeen materiaal (sterretjes) en deze methoden worden dus voornamelijk gebruikt voor modelsoorten. TRAP is vooral geschikt voor plantenwetenschappelijke studies waarbij celwand (CW) of mechanische signalering en celsoorten die moeilijk te bevrijden zijn uit hun CW-matrix. B. Gedetailleerde wet-lab stappen van de TRAP procedure: Zaailingen uitdrukken GFP-gelabeld ribosomal eiwit in verschillende celtypes (bijvoorbeeld wortel endodermis) worden geteeld op petrischaaltjes gedurende zeven dagen en wortelmateriaal geoogst door snap bevriezing. Een totaal RNA controlemonster wordt verzameld uit het gehomogeniseerde ruwe extract voordat pelleting het puin via centrifugeren. Magnetische anti-GFP kralen worden toegevoegd aan het gewiste extract om immunoneerslag uit te voeren. Na incubatie en drie wasstappen wordt het polysome-geassocieerde RNA (TRAP/polysome RNA) rechtstreeks verkregen via fenol-chloroform extractie. LCM: lasercapture microdissectie, FACS/FANS: fluorescentie-geactiveerde cel/nucleaire sortering, APEX-seq: methode gebaseerd op ontworpen ascorbate peroxidase, INTACT: isolatie van kernen die zijn getagd in specifieke celtypen, SLAM-ITseq: thiol(SH)-gekoppelde alkylation voor de metabole sequencing van RNA in weefsel, Flura-seq: fluorourcila-label RNA-sequencing (Gemaakt met Biorender.com) Klik hier om een grotere versie van dit cijfer te bekijken.
Het doel van dit artikel is om een gedetailleerde beschrijving van de TRAP-methode te geven, om kritieke stappen te markeren en om richtlijnen te bieden voor een mogelijke methode voor het voorbereiden van bibliotheken.
Een generiek TRAP-experiment zal hoofdzakelijk bestaan uit de volgende stappen (zie ook figuur 1B):(1) Bereiding van plantaardig materiaal, met inbegrip van het klonen van ribosoom-tagging constructie, transgene lijnproductie en -selectie, het kweken en ophopen van zaden, sterilisatie en beplating, en stresstoepassing/behandeling (optioneel) en weefseloogsten; (2) immunozuivering, met inbegrip van weefselhomogenisatie en clearing van het ruwe extract, de wassen van de kraal en de immunozuivering, en de wasstappen; (3) RNA-extractie en kwaliteitsbeoordeling; en (4) bibliotheekvoorbereiding.
De Arabidopsis wortel is een modelsysteem om plantontwikkeling te bestuderen sinds de introductie als een model plant37,38. Hier wordt de toepassing van TRAP getoond in de context van de ontwikkeling van de wortel van planten. Bij planten is de opbouw van het gehele wortelstelsel afhankelijk van de uitvoering van dit programma en is daarom zeer belangrijk voor de overleving van het organisme39. In Arabidopsiszijn laterale wortels afkomstig van pericycleweefsel dat naast xylemvaten ligt en daarom xylempaalpericycle (XPP; zie figuur 2C)40. Sommige XPP-cellen, die zich diep in de wortel bevinden, verwerven een identiteit van de stichterscel en beginnen zich, na een lokale hormonale trigger, te vermenigvuldigen door anticlinally te zwellen en te delen41. Echter, als gevolg van de aanwezigheid van een stijve celwand matrix, dit proces oefent mechanische stress op de omliggende weefsels. Met name de bovenliggende endodermis wordt beïnvloed, omdat deze in de weg staat van de laterale wortelgroeias42,43,44. Inderdaad, de nieuw gevormde primordium zal moeten groeien door de bovenliggende endodermis cel (Figuur 2C2), terwijl cortex en opperhuid cellen zijn gewoon opzij geschoven voor de primordium om eindelijk te voorschijn45,46. Recent werk in ons lab heeft aangetoond dat de endodermis actief bijdraagt aan de proliferatie in de pericycle. Gerichte blokkering van endodermale hormonale signalering is voldoende om zelfs de allereerste divisie in de XPP-cellen47te remmen. Vandaar, pericycle-endodermis communicatie vormt een zeer vroege checkpoint voor laterale wortel ontwikkeling in Arabidopsis. Het is echter niet bekend hoe deze crosstalk wordt uitgevoerd. Om dit mysterie te ontrafelen, kozen we voor de TRAP-seq-benadering om XPP- en dodermale cellen te targeten. Om te verrijken voor cellen in het laterale wortelprogramma, bootsten we de hormonale trigger na door een auxine analoog (1-naphthaleneacetisch zuur, NAA)48exogenly toe te passen, waardoor tegelijkertijd de eerste fase van laterale wortelvorming tijdelijk kon worden opgelost.
