Poussé par l’application croissante des techniques de séquençage de nouvelle génération, la résolution spatiale en biologie du développement pourrait être augmentée. Les études contemporaines visent à disséquer les tissus vers le bas pour les types de cellules spécialisées, si ce n’est le niveau unicellulaire1,2,3,4. À cette fin, une pléthore de différentes méthodes a été conçue au cours des cinquante dernières années (voir figure 1A)5,6,7,8,9,10,11,12,13,14,15.
De nombreux outils en science végétale ont été des adaptations de techniques qui ont été pionnières dans la recherche animale. Ce n’est pas le cas pour la méthode que nous introduisons en détail ici. En 2005, doté d’un solide fond de traduction de protéines, le Laboratoire Bailey-Serres a entrepris de concevoir des protéines ribosomales pour la purification ultérieure de l’affinité16. Ainsi, ils pourraient éviter le profilage polysome long et laborieux, qui est basé sur l’ultracentrifugation avec un gradient de saccharose et a été employé pour évaluer la traduction des ribosomes depuis les années 196017,18. La méthode a depuis été appelée purification d’affinité ribosome translationnelle (TRAP)16. Après des études réussies de traduction dans les plantes, Heiman et autres ont adapté TRAP pour les animaux19 et d’autres ont étendu son application à la levure20, Drosophila21, Xenopus22 et le poisson zèbre23,24.
Bien que la modification génétique du système modèle soit une condition préalable pour TRAP, qui limite son application aux espèces qui se prêtent à la transformation génétique, on peut simultanément exploiter cette objection pour cibler des sous-ensembles de cellules qui sont d’intérêt particulier et autrement extrêmement difficiles à isoler du tissu intact/organe25 (p. ex., les cellules dendritiques fortement ramifiées dans un cerveau de souris ou hyphes fongiques dans le tissu végétal infecté). Dans les plantes, toutes les cellules sont maintenues en place par des parois cellulaires qui forment la base du squelette hydrostatique26. Pour libérer une cellule végétale de cette matrice, les scientifiques ont soit physiquement coupé la cellule de son tissu environnant par microdissection de capture laser (LCM)27 ou effectué la digestion enzymatique des parois cellulaires28. Parmi ces dernières cellules, ce que l’on appelle les protoplastes, la population d’intérêt est étiquetée fluorescentement et peut être séparée par le tri cellulaire activé par fluorescence (FACS)7. LCM nécessite généralement un échantillon à fixer et intégré dans la cire, ce qui finit par détériorer la qualité de son ARN29. Les méthodes à base de FACS produisent de l’ARN de haute qualité, mais le processus de protoplasting lui-même introduit des différences dans l’expressiongénique 30 et les tissus avec des parois cellulaires secondaires modifiées et épaisses sont notoirement difficiles à traiter. En outre, de nombreux processus de développement dans les plantes sont supposés s’appuyer sur des signaux mécaniquement transmis et donc l’intégrité de la paroi cellulaire est d’une importance primordiale31. Deux méthodes, qui utilisent un raccourci pour contourner l’isolement cellulaire en opérant au niveau des nucléii, sont le tri nucléaire activé par la fluorescence (FANS) et l’isolement des noyaux marqués dans des types cellulaires spécifiques (INTACT). Comme dans TRAP, ils utilisent des promoteurs spécifiques au type de cellule pour marquer les noyaux, qui se sont ensuite enrichis par tri ou tirer vers le bas, respectivement8,15. Un défi majeur pour toutes ces approches est d’obtenir suffisamment de matériel d’ARN à partir de sous-ensembles de cellules dans un tissu. Comme TRAP ne capture qu’une fraction des ARN cellulaires, la collecte d’échantillons est un goulot d’étranglement considérable. Par conséquent, des protocoles de préparation de bibliothèque particulièrement sensibles sont nécessaires pour produire des données de haute qualité à partir de faibles quantités d’entrée.
Depuis sa création, TRAP a été utilisé en combinaison avec des microrésions d’ADN ou, comme les coûts de séquençage ont chuté de manière significative ces dernières années, RNA-seq10,32,33. Une multitude de questions de recherche a déjà été élucidée tel qu’il a été examiné dans Sablok et coll.34. Nous sommes convaincus que d’autres rapports suivront dans les années à venir, car la technique est très polyvalente lorsqu’elle combine différents promoteurs pour cibler des types de cellules spécifiques. Finalement, cela se fera même d’une manière inductible, et peut être combiné avec sonder la réaction de la plante à de nombreux facteurs de stress biotiques et abiotiques. En outre, lorsque des lignées transgéniques stables ne sont pas disponibles, les systèmes d’expression des racines poilues ont également été utilisés avec succès pour effectuer TRAP dans la tomate et medicago35,36.
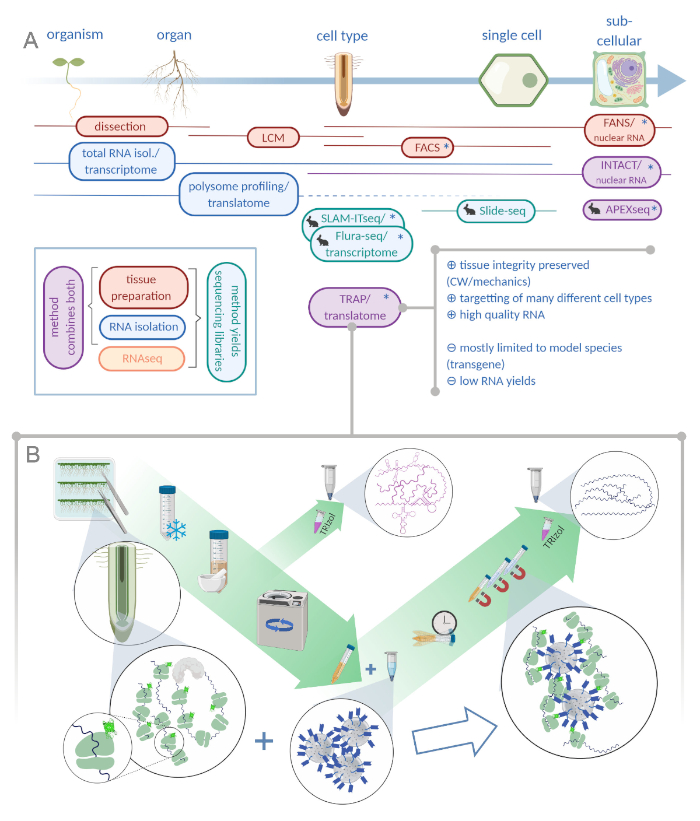
Figure 1 : La traduction de la purification de l’affinité ribosome (TRAP) complète le portefeuille d’analyse « omics ». R. L’augmentation des niveaux de précision analytique, jusqu’à la résolution unicellulaire ou même subcellulaire peut être obtenue par une pléthore de méthodes ou de combinaisons de celui-ci. Le programme donne un aperçu des outils actuellement disponibles dans le domaine des plantes et des animaux. La collecte de tissus à la résolution cellulaire peut être obtenue par des protocoles comme LCM ou FACS, qui sont ensuite couplés à un transcriptome standard ou à une analyse de profilage/traduction polysome. TRAP et INTACT intègrent à la fois la capture des tissus et l’isolement de l’ARN car ils sont basés sur l’épitope-marquage. Cependant, INTACT n’échantillonne que les noyaux cellulaires et constitue, par conséquent, un cas particulier d’analyse de transcriptome. Une petite icône de lapin marque des méthodes nouvellement développées dans le domaine animal : Alors que SLAM-ITseq et Flura-seq s’appuient sur la ciblage métabolique des ARN naissants avec des bases d’uracil modifiées dans les cellules exprimant l’enzyme permissive, Slide-seq utilise une diapositive en verre enduit avec des codes-barres d’ADN qui fournissent des informations de position dans la gamme cellulaire. Une approche d’étiquetage de proximité est suivie dans APEX-seq pour échantillonner les ARN dans des compartiments subcellulaires spécifiques. Notamment, une résolution accrue nécessite souvent la production de matériaux transgéniques (astérisques) et ces méthodes sont donc principalement utilisées pour les espèces modèles. TRAP est particulièrement adapté pour les études scientifiques végétales impliquant la paroi cellulaire (CW) ou la signalisation mécanique ainsi que les espèces cellulaires qui sont difficiles à libérer de leur matrice CW. B. Les étapes détaillées de la procédure TRAP : Les semis exprimant des protéines ribosomal étiquetées GFP dans des types cellulaires distincts (p. ex. endodermis de racine) sont cultivés sur des plats Petri pendant sept jours et des racines récoltées par congélation rapide. Un échantillon total de contrôle de l’ARN est prélevé sur l’extrait brut homogénéisé avant de pelleter les débris par centrifugation. Des perles magnétiques anti-GFP sont ajoutées à l’extrait effacé pour effectuer l’immunoprécipitation. Après l’incubation et trois étapes de lavage, l’ARN polysome-associé (TRAP/ARN polysome) est directement obtenu par extraction de phénol-chloroforme. LCM : microdissection de capture laser, FACS/FANS : tri cellulaire/nucléaire activé par fluorescence, APEX-seq: méthode basée sur l’ascorbate peroxidase, INTACT: isolation des noyaux étiquetés dans des types cellulaires spécifiques, SLAM-ITseq: thiol (SH) alkylation liée à la séquence métabolique de l’ARN dans les tissus, Flura-seq: fluorouracil-étiqueté RNA séquençage (Créé avec Biorender.com) S’il vous plaît cliquez ici pour voir une version plus grande de cette figure.
Le but de cet article est de fournir une description détaillée de la méthode TRAP, de mettre en évidence les étapes critiques et de fournir des conseils pour une méthode de préparation possible de la bibliothèque.
Une expérience TRAP générique consistera essentiellement en les étapes suivantes (voir aussi la figure 1B) : (1) Préparation du matériel végétal, y compris le clonage de la construction de ribosome-marquage, la production et la sélection transgéniques de lignées, la culture et l’encombrement des graines, la stérilisation et le placage, et l’application/traitement du stress (facultatif) et la récolte des tissus; (2) l’immunopurification, y compris l’homogénéisation des tissus et le dégagement de l’extrait brut, le lavage et l’immunopurification des perles, et les étapes de lavage; (3) Évaluation de la qualité de l’ARN; et (4) la préparation de la bibliothèque.
La racine Arabidopsis a été un système modèle pour étudier le développement des plantes depuis son introduction comme une plante modèle37,38. Ici, l’application de TRAP est présentée dans le contexte du développement latéral des racines végétales. Dans les plantes, l’accumulation de l’ensemble du système racinaire repose sur l’exécution de ce programme et est donc très importante pour la survie de l’organisme39. Dans Arabidopsis, les racines latérales proviennent de tissu péricycle qui réside à côté des vaisseaux xylèmes et est donc appelé pericycle pôle xylème (XPP; voir figure 2C)40. Certaines cellules XPP, qui sont situées profondément à l’intérieur de la racine, acquièrent une identité cellulaire fondatrice et, sur un déclencheur hormonal local, commencent à proliférer en gonfleant et en divisant anticlinally41. Cependant, en raison de la présence d’une matrice rigide de mur cellulaire, ce processus exerce le stress mécanique sur les tissus environnants. En particulier, l’endoderme overlying est affecté, car il est dans la voie de l’axe de croissance latérale des racines42,43,44. En effet, le primordium nouvellement formé devra se développer à travers la cellule endodermis overlying (figure 2C2) tandis que les cellules cortex et épiderme sont juste repoussées pour que le primordium émerge enfin45,46. Des travaux récents dans notre laboratoire ont montré que l’endoderme contribue activement à la prolifération du péricycle. Le blocage ciblé de la signalisation hormonale endodermique est suffisant pour inhiber même la toute première division dans les cellules XPP47. Par conséquent, la communication péricycle-endodermis constitue un point de contrôle très précoce pour le développement latéral des racines dans Arabidopsis. On ne sait cependant pas comment cette traque est effectuée. Pour percer ce mystère, nous avons choisi l’approche TRAP-seq pour cibler XPP et les cellules endodermiques. Pour enrichir pour les cellules dans le programme de racine latérale, nous avons imité le déclencheur hormonal en appliquant exogènement un analogue auxin (acide à laphthalèneacécéétique, NAA)48, qui en même temps a permis de résoudre temporellement la phase initiale de la formation latérale de racine.
