As described in the diagram (Figure 1A), the session starts with pre-exposure (Day 1), followed by fear conditioning (Days 2 and 3), and then either extinction or retrieval (Day 4).
Presentations of the SCS in the pre-exposure (Day 1) session did not elicit flight or freezing response in the mice (Figure 2A-2B). Behavioral analysis during conditioning (Days 2 and 3) revealed that the tone component of the SCS significantly enhanced freezing compared to freezing during the pre-SCS (Figure 2B,2E). Flight scores changed significantly across sessions (Day 1 to Day 3, n = 20; Figure 2A). Mice showed higher speed and more jumps, and thus greater flight scores, to the white noise cue compared to tone (Figure 2C-2D). Mice showed a clear transition of defensive behavior–exhibiting lower flight scores during the tone followed by higher flight scores during white noise (Figure 2F) and vice-versa for freezing responses (Figure 2G).
To test for the effect of threat proximity and context on conditioned flight, mice were split into two groups: one group underwent extinction training in the conditioning context (Figure 3A-3B), and another group was tested for fear memory recall by exposing them to the SCS in a neutral context (Figure 3C-3D). Mice subjected to the 16 trials of extinction training showed rapid extinction of conditioned flight (n = 12). Flight scores during the first block of four trials were higher during white noise as compared to the tone (Figure 3A). Flight behavior was no longer elicited by either cue at the end of the extinction session. There was an overall decrease in tone-induced freezing and an increase in white noise-mediated freezing during the extinction session. Freezing for the first block of four trials was significantly higher to the tone compared to the white noise (Figure 3B). This suggests imminence of the threat is vital for the flight response.
The flight response was diminished in a context-dependent manner. Exposure to the white noise in the neutral context did not elicit flight (n = 8). Instead, white noise presentations in the neutral context elicited freezing responses which were higher than those elicited by the tone (Figure 3C-3D). This demonstrates the importance of context in modulating defensive responding.
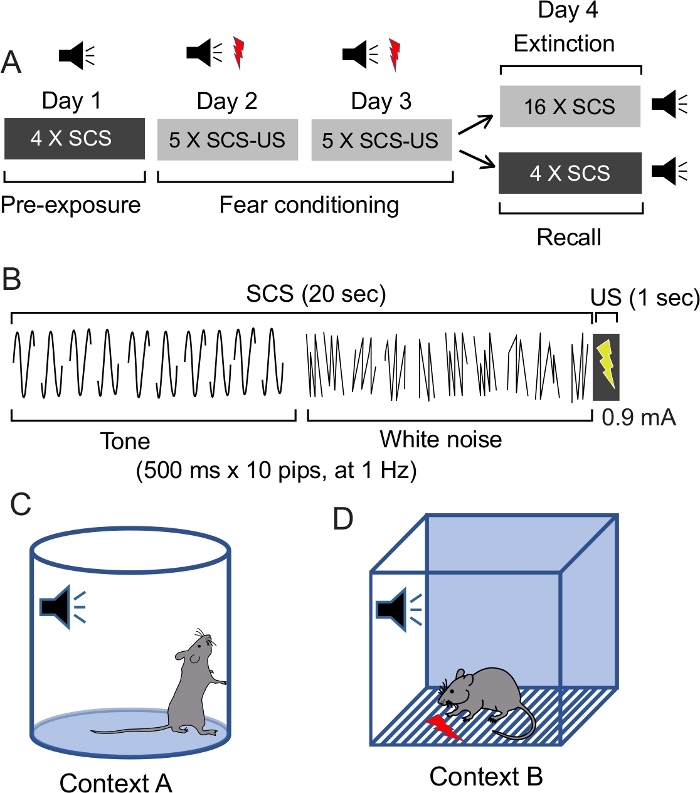
Figure 1: Study design to assess freezing and flight paradigm.
A) Diagram of the behavioral sessions. B) Diagram detailing the composition of the serial compound stimulus (SCS), as well as the timing of the US. C) Context A – served as a neutral context, and used during pre-exposure and recall sessions. D) Context B – used for fear conditioning. This figure has been modified from Fadok et al. 2017. Please click here to view a larger version of this figure.
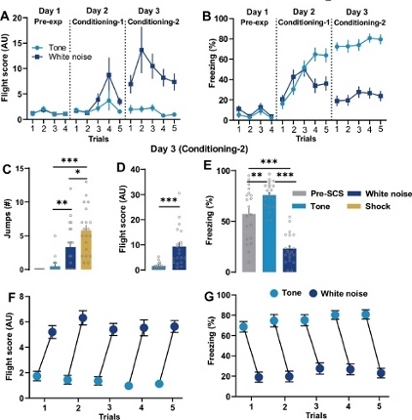
Figure 2: Conditioned flight response.
A) Comparison of average trial-wise flight scores (n = 20) following presentation of the tone and white noise across Days 1-3. A significant change in the flight scores across sessions has been noted (Day 1 to Day 3; two-way repeated measures ANOVA, cue × trial interaction, F (13, 266) = 5.795; P<0.0001). Post-hoc Bonferroni’s multiple comparison test reveals a significant difference between tone and white noise induced flight scores at fear conditioning Day 1 (trial 4, P < 0.05) and Day 2 (trials 2-5, P < 0.001). B) Comparison of average trial-wise % freezing during the tone and white noise periods across Days 1-3. Note a statistically significant change in % freezing across the sessions (Day 1 to Day 3, n = 20; two-way repeated-measures ANOVA, cue × trial interaction, F (13, 266) = 20.81; P < 0.001; Figure 2B). Post-hoc Bonferroni’s multiple comparison test reveals a significant difference between tone and white noise induced freezing at fear conditioning Day 1 (trial 4 and 5, P < 0.001) and Day 2 (all trials, P < 0.001). C) Comparison of number of jump escape responses during the pre-SCS, tone, white noise, and shock periods on Day 3. One-way ANOVA followed by Bonferroni's multiple comparisons test showed that escape jumps were significantly higher during white noise and shock as compared to tone period (P < 0.01 and P < 0.001, respectively). D) Comparison of flight scores during the presentation of tone and white noise on Day 3. Note a significantly higher flight scores on Day 3 during white noise period (P < 0.001, Wilcoxon matched-pairs signed-rank test). E) Comparison of % freezing during the pre-SCS, tone, and white noise on Day 3. Freezing behavior on Day 3 reveals significant effect of tone and white noise (one-way repeated-measures ANOVA, F = 56.82, P<0.01). Bonferroni's multiple comparisons test showed that presentation of tone significantly increases % freezing vs pre-SCS duration (P < 0.01), whereas % freezing was significantly reduced as compared pre-SCS and tone durations (both P < 0.001). The representative trial-wise data shows transitions of flight (F) and freezing (G) behavior following the presentation of tone and white noise in the mouse on Day 3. The represented values are means ± SEM. *P<0.05, **P<0.01, ***P <0.001. Pre-exp, Pre-exposure. Panels A-E are modified from Fadok et al., 2017. Please click here to view a larger version of this figure.
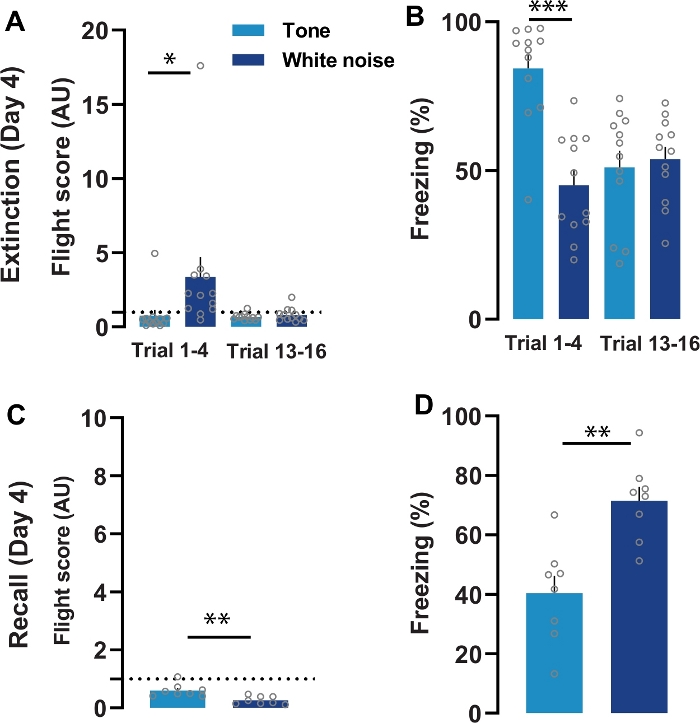
Figure 3: Extinction and recall following flight conditioning (Day 4).
A) Comparison of flight scores during extinction training showed rapid extinction of conditioned flight (n = 12; 16 trials, two-way repeated-measures ANOVA, cue × trial interaction, F(15,165) = 3.05, P < 0.01). Flight scores from first block of four trials (trial 1-4) of extinction observed significantly higher for white noise as compared to the tone (P < 0.05, Wilcoxon matched-pairs signed-rank test). B) Comparison of freezing showed a statistically significant effect on freezing (%) following white noise (n = 12; 16 trials, two-way repeated-measures ANOVA, cue × trial interaction, F(15,165) = 3.55, P < 0.01). The freezing for the first block of four trials (trial 1-4) during extinction found to be significantly lower during white noise period as compared to the tone (Paired t-test, P < 0.01). C) Changes in the context significantly affect the flight scores (n = 8; 4 trials, two-way repeated-measures ANOVA, cue × trial interaction, F(1,7) = 27.44, P < 0.01). Flight scores significantly reduced during white noise as compared to the tone period in the neutral context (two-tailed paired t-test, P < 0.01) D). Freezing responses across trials during retrieval were also significant (n = 8, 4 trials, two-way repeated-measures ANOVA, effect of cue F(1,7) = 27.67, P < 0.01). Exposure of WN in neutral context significantly increased the freezing responses as compared to the tone (two-tailed paired t-test, P < 0.001). The represented values are means ± SEM. *P<0.05, **P<0.01, ***P <0.001. Panels A-D are modified from Fadok et al. 2017. Please click here to view a larger version of this figure.
