Dinâmica de sondagem de matéria macia com nêutrons
Investigar as propriedades dinâmicas de proteínas e peptídeos é uma parte importante da pesquisa biofísica, e muitos métodos bem desenvolvidos existem hoje para acessar uma ampla gama de paisagens energéticas1. Relacionar a dinâmica experimentalmente revelada das proteínas à sua função biológica é uma tarefa muito mais difícil, exigindo modelos matemáticos complexos e simulações dinâmicas auxiliadas por computador. A importância da espectroscopia de nêutrons para a análise de movimentos proteicos tem sido enfatizada em diversos estudos bem recebidos e amplamente reconhecidos 1,2,3,4,5. Antes de explorar a paisagem energética diversificada da dinâmica interna de proteínas, uma breve visão geral dos processos dinâmicos em matéria suave e como os nêutrons podem acessá-los é necessária.
A sensibilidade dos nêutrons à configuração isotópica e o tipo de interações que exibem com matéria macia faz da dispersão de nêutrons uma das técnicas de investigação mais versáteis6. Há um amplo espectro de escalas de comprimento de correlação e tempos de correlação que os nêutrons podem acessar, desde excitações nucleares e vibrações atômicas até movimentos coletivos e processos de relaxamento lentos, como rotações isotrópicas e movimentos difusivos. Ao investigar os nêutrons dispersos para sua transferência de energia, três interações principais podem ser distinguidas: a dispersão elástica, na qual não há troca de energia entre nêutrons e partículas de entrada na amostra; a dispersão inelástica, com uma grande troca de energia quantificável entre nêutrons e partículas; e o caso peculiar de dispersão quase elástica que designa uma transferência de energia muito pequena em comparação com o incidente de energia de nêutrons 1,7. Essas interações dão informações precisas sobre o material investigado e formam a base teórica de uma ampla variedade de técnicas de dispersão de nêutrons.
Em dispersão elástica, o detector registra as direções dos nêutrons como um padrão de difração, que mostra a posição dos átomos amostrais em relação um ao outro. Informações sobre as correlações das posições atômicas são adquiridas (ou seja, intensidade integrada S(Q) relativa à transferência de impulso Q, que diz respeito apenas à informação estrutural). Este princípio forma a base da difração de nêutrons8.
A complexidade surge quando a transferência de energia não é mais zero devido a excitações e flutuações internas no material amostral. Isso forma a base da espectroscopia de nêutrons, na qual os nêutrons dispersos são investigados em função tanto da transferência de energia E quanto da transferência de impulso Q. Informações dinâmicas e estruturais são obtidas. A espectroscopia de nêutrons mede a mesma intensidade integrada S(Q) para transferência de energia (ou seja, mudança de velocidade dos nêutrons devido à dispersão amostral, S(Q,ω) = S(Q, E), que também é referido como o fator de estrutura dinâmica)9.
Para calcular a dispersão de um material, é mais adequado usar a função de correlação do par 7,10. No caso da difração, a função de correlação do par estático G(r) dá a probabilidade de encontrar o centro de uma partícula a uma determinada distância r do centro de outra partícula. A espectroscopia generaliza a função de correlação do par estático e inclui energia/frequência/tempo na equação de dispersão. A função de correlação de par G(r) torna-se uma função do tempo G(r, t), que pode ser decomposta em uma função de correlação de par de átomos distinta GD(r, t), e uma função de auto-correlação GS(r, t). Estes descrevem dois tipos de correlações: movimentos correlacionados por pares de átomos que regem a dispersão coerente, e a auto-correlação que rege a dispersão incoerente10.
A dispersão coerente é a dispersão da “média” e depende da fase relativa das ondas dispersas. No regime de dispersão de pequenos ângulos, as ondas de nêutrons dispersas de diferentes centros de dispersão (diferentes átomos) interferem construtivamente (têm fases semelhantes), e o movimento coletivo dos átomos é observado com forte aumento de intensidade. A dispersão coerente descreve essencialmente a dispersão de um único nêutron de todos os núcleos da amostra10.
Quando não ocorre interferência construtiva entre as ondas de nêutrons dispersas de diferentes centros, um único átomo é seguido no tempo, e a auto-correlação entre a posição do átomo no momento t = 0 e o mesmo átomo no momento em que t é observado. Assim, as informações sobre as posições relativas dos átomos são perdidas, e o foco está apenas nas flutuações locais. A dispersão das flutuações locais rege a dispersão incoerente. A dispersão incoerente é isotrópica, contribui para o sinal de fundo e degrada o sinal-para-ruído10,11.
Combinando todos os processos acima, distinguimos quatro grandes processos de dispersão de nêutrons10: (1) elástico coerente (mede as correlações das posições atômicas), (2) inelástico coerente (mede movimentos coletivos de átomos), (3) elástico incoerente (contribui para o fundo, reduz a intensidade de dispersão pelo fator Debye-Waller (DWF) e mede o fator de estrutura incoerente elástico (EISF), descrevendo a geometria dos movimentos difusivos na geometria confinada, e (4) incoerentes inelesses (mede dinâmica de átomo único e auto correlação).
Os processos dinâmicos que os nêutrons podem acessar na biologia vão desde o amortecimento de vibrações atômicas e moleculares de baixa frequência, a interação de moléculas de solventes com bio-superfícies e processos de difusão na camada de hidratação de macromoléculas e geometria confinada, até movimentos translacionais, rotacionais e difusivos de curto alcance, e domínios proteicos e movimentos alotéricos1 . A ampla diversidade de métodos e instrumentos de nêutrons para medir a dinâmica proteica baseia-se na forma como a acromatização do feixe de nêutrons incidente ou saída é alcançada e como a análise energética dos nêutrons dispersos é realizada. De eixo triplo a espectrômetros de retorno e spin-echo, pode-se explorar processos dinâmicos com tempos característicos entre 1 x 10-14 s e 1 x 10-6 s (femtosegundos a microsegundos)12.
Oak Ridge National Laboratory, com suas duas renomadas fontes de nêutrons, a Fonte de Nêutrons Spallation – SNS13 e o Reator de Fluxo de Alto Isótopo – HFIR14, tem uma das melhores suítes de espectrômetros para investigar dinâmicas em bioquímicos. Alguns dos exemplos mais eloquentes incluem o uso do espectrômetro de helicóptero de nêutrons frio (CNCS) no SNS15 para investigar a perturbação dinâmica da água de hidratação em torno da proteína fluorescente verde na solução16 ou as vibrações coletivas sub-picosegundos de várias proteínas17. Um problema recorrente das investigações inelásticas de dispersão de nêutrons é que alguns processos biológicos são muito lentos para serem observados. Sem configurações extremas que levam a uma enorme perda de intensidade de nêutrons, os espectrômetros de tempo de voo são limitados à resolução de energia de 10 μeV, correspondendo a uma escala de tempo máxima de ~200 ps10,11. Isso não é suficiente para observar movimentos em larga escala em proteínas. Portanto, instrumentos com maior resolução de energia, como os espectrômetros de backscattering, são frequentemente necessários. A combinação das técnicas de tempo de voo e backscattering tem se mostrado poderosa para investigar a mudança na dinâmica interna do Cytochrome P450cam (CYP101), uma enzima que catalisa a cânfora de hidroxiagem18.
A difusividade microscópica medida pelo espectrômetro de backscattering no SNS-BASE19 foi surpreendentemente bem definida e poderia ser separada na difusividade da água (hidratação, citoplasmático e água a granel) e a difusividade dos constituintes celulares em flatworms planárias, o primeiro animal vivo a ser estudado pela dispersão de nêutrons20 . Backscattering é uma técnica espectroscópica de alta resolução, mas também é limitada a vários μeV = vários nanossegundos, enquanto a dinâmica lenta em biomateriais também se manifesta como o tempo de sobrevivência da correlação entre posição atômica ou orientações de spin (por exemplo, processos de relaxamento, que acontecem regularmente na faixa de tempo de dez a centenas de nanossegundos).
A espectroscopia de eco de giro de nêutrons (NSE) é a única técnica de dispersão de nêutrons a atingir tal alta resolução. Ao contrário de outras técnicas de nêutrons, a NSE não requer acromatização do feixe, pois usa a fase mecânica quântica dos nêutrons, que são seus momentos magnéticos. A manipulação de momentos magnéticos permite o uso de uma ampla distribuição de comprimento de onda de feixe de nêutrons, enquanto a técnica é sensível a pequenas mudanças de velocidade de nêutrons na ordem de 1 x 10-4. A NSE tem sido usada com sucesso para investigar a lenta dinâmica das proteínas em solução para muitas proteínas. Entre esses muitos estudos pioneiros, reconhecemos o estudo da flexibilidade segmental da imunoglobulinasuína 21; os movimentos de domínio acoplado na polimerase Taq22; os movimentos de domínio no tetramer do álcool levedura desidrogenase23; a alteração da conformação na quinase fosfoglicerato sobre a ligação do substrato3; a ativação dos movimentos de domínio e a propagação dinâmica de sinais aluséricos na proteína de cofactor regulatório de câmbio Na+/H+ 1 (NHERF1) 4,24,25; a dinâmica de um estado compacto de redução de íons mercuricos26; e a difusão da hemoglobina em glóbulos vermelhos27. Dois estudos mais recentes em dinâmica proteica expuseram a flexibilidade do anticorpo humano Imunoglobulina G (IgG) como uma mola entropica28 e as características da contribuição do solvente para a dinâmica da proteína básica de mielina intrinsecamente desordenada (MBP)5.
O presente artigo explica os princípios básicos da NSE, os múltiplos métodos preparatórios recomendados para uma investigação minuciosa da dinâmica proteica, bem como a metodologia e o protocolo experimental para aquisição de dados NSE no espectrômetro NSE na SNS, SNS-NSE. O protocolo caracteriza duas proteínas: IgG, uma proteína de anticorpos humanos regular, e a proteína intrinsecamente desordenada MBP. As implicações biofísicas, a relevância da pesquisa dos exemplos e as limitações da técnica são discutidas brevemente.
Espectroscopia NSE, o método para medições dinâmicas lentas
NSE é uma técnica polarizada que usa o tempo de voo de nêutrons para medir a troca de energia (perda de polarização) devido à interação quase elástica entre nêutrons e átomos em uma amostra. No núcleo da espectroscopia NSE estão dois princípios básicos: (1) a capacidade do giro de nêutrons para precess no campo magnético com uma frequência proporcional à força 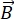 magnética , ou seja, a frequência larmor29, e (b) o spin-echo ou Hann echo, representando a manipulação e refoco do sinal de polarização ao aplicar uma série de pulsos de radiofrequência30.
magnética , ou seja, a frequência larmor29, e (b) o spin-echo ou Hann echo, representando a manipulação e refoco do sinal de polarização ao aplicar uma série de pulsos de radiofrequência30.
O básico do processo NSE pode ser resumido em algumas etapas simples 6,11 usando a Figura 1. (1) O feixe de nêutrons produzido pela fonte (posição 1) é polarizado (posição 2), guiado e transportado (posição 3), e chega à entrada do espectrômetro NSE, onde é girado por 90° pela primeira nadadeira pi-half (posição 4). (2) O feixe polarizado (por exemplo, momentos magnéticos de nêutrons) torna-se perpendicular às linhas de campo magnético do primeiro ímã (primeira zona de pré-cessão, posição 5) e começa a precess. (3) No final do ímã, os giros de nêutrons acumulam um certo ângulo de pré-cessão proporcional à força do campo magnético e ao tempo de voo gasto no interior (basicamente inversamente proporcional à velocidade de nêutrons). As velocidades individuais de nêutrons são codificadas dentro de seu ângulo de precessão no final da primeira zona de precessão. (4) Perto da posição da amostra, o pi-flipper (posição 6) inverte a orientação da rotação em 180°, alterando o sinal do ângulo de precessão. (5) Os nêutrons interagem com as moléculas da amostra (posição 7) e ficam dispersos. (6) Os nêutrons dispersos entram e precess na segunda zona de pré-cessão (posição 8), mas tornam-se orientados para a inversão. (7) Outra nadadeira pi-half (posição 9) é usada para girar a orientação do giro do perpendicular à direção horizontal. Isso impedirá a precessão, traduzindo o ângulo de precessão φ em polarização proporcional à cos(φ). (8) O analisador (posição 10) seleciona os nêutrons com base em uma orientação. Se a interação com a amostra for elástica, a velocidade do nêutron não mudará. Os nêutrons passarão um tempo idêntico voando na primeira e segunda zonas de pré-cessão, e os ângulos acumulados de precessão estão totalmente recuperados. A polarização completa é restaurada no detector (posição 11) como um eco da polarização original (ou seja, spin-echo). (9) No entanto, em NSE, a dispersão é quase elástica, de modo que uma pequena troca de energia entre nêutrons e moléculas de amostras leva a diferentes velocidades de nêutrons após a dispersão pela amostra. Devido às diferentes velocidades, os nêutrons passarão um tempo adicional voando pela segunda zona de pré-cessão e não terão recuperado adequadamente seu ângulo de precessão. Uma polarização parcial é recuperada no detector, e a perda de polarização devido ao relaxamento do giro é proporcional à transformação cos-Fourier da função espectral S(Q, ω), a função de dispersão intermediária F(Q, t). (10) O parâmetro de tempo da função F(Q, t) é proporcional à força do campo magnético de pré-cessão. A varredura da perda de polarização em função da força do campo magnético rende, portanto, uma função de relaxamento que depende dos processos dinâmicos dentro da amostra.

Figura 1: Fotografia do espectrômetro NSE no SNS (SNS-NSE) e esquema de caminho de mosca de nêutrons com os componentes funcionais mais importantes. Da direita para a esquerda: 1 = fonte de nêutrons; 2 = sistema de obturadores-bender-bender-polarizador-secundário; 3 = guias de transporte de vigas; 4 = flipper pi/2 para o primeiro giro de 90°; 5 = primeira zona de pré-cessão; 6 = flipper pi para giro de 180°; 7 = área amostral e ambiente amostral (aqui, mostra-se o crio-forno); 8 = segunda zona de pré-cessão; 9 = pi/2 flipper para o segundo giro de 90°; 10 = analisador; 11 = detector. (Observe que as porções de 3, bem como 2 e 1 , estão situadas atrás da parede azul dentro da blindagem; os helicópteros são substituídos por um seletor de velocidade para NSE baseado em reator). Clique aqui para ver uma versão maior desta figura.
