Dynamique de sondage de la matière molle avec des neutrons
L’étude des propriétés dynamiques des protéines et des peptides est une partie importante de la recherche biophysique, et de nombreuses méthodes bien développées existent aujourd’hui pour accéder à un large éventail de paysages énergétiques1. Relier la dynamique révélée expérimentalement des protéines à leur fonction biologique est une tâche beaucoup plus difficile, nécessitant des modèles mathématiques complexes et des simulations dynamiques assistées par ordinateur. L’importance de la spectroscopie neutronique pour l’analyse des mouvements des protéines a été soulignée dans plusieurs études bien reçues et largement reconnues 1,2,3,4,5. Avant d’explorer le paysage énergétique diversifié de la dynamique interne des protéines, un bref aperçu des processus dynamiques dans la matière molle et de la façon dont les neutrons peuvent y accéder est nécessaire.
La sensibilité des neutrons à la configuration isotopique et le type d’interactions qu’ils présentent avec la matière molle font de la diffusion des neutrons l’une des techniques d’investigation les plus polyvalentes6. Il existe un large spectre d’échelles de longueur de corrélation et de temps de corrélation auxquels les neutrons peuvent accéder, des excitations nucléaires et des vibrations atomiques aux mouvements collectifs et aux processus de relaxation lents comme les rotations isotropes et les mouvements diffusifs. Lors de l’étude des neutrons diffusés pour leur transfert d’énergie, trois interactions principales peuvent être distinguées: la diffusion élastique, dans laquelle il n’y a pas d’échange d’énergie entre le neutron entrant et la particule dans l’échantillon; la diffusion inélastique, avec un grand échange d’énergie quantifiable entre le neutron et la particule; et le cas particulier de la diffusion quasi-élastique qui désigne un très petit transfert d’énergie par rapport à l’énergie neutronique incidente 1,7. Ces interactions donnent des informations précises sur le matériau étudié et constituent la base théorique d’une grande variété de techniques de diffusion des neutrons.
En diffusion élastique, le détecteur enregistre les directions des neutrons sous la forme d’un motif de diffraction, qui montre la position des atomes de l’échantillon les uns par rapport aux autres. Des informations sur les corrélations des positions atomiques sont acquises (c’est-à-dire l’intensité intégrée S(Q) concernant le transfert de quantité de mouvement Q, qui se rapporte uniquement à l’information structurelle). Ce principe constitue la base de la diffractionneutronique 8.
La complexité survient lorsque le transfert d’énergie n’est plus nul en raison des excitations et des fluctuations internes du matériau de l’échantillon. Cela constitue la base de la spectroscopie neutronique, dans laquelle les neutrons diffusés sont étudiés en fonction du transfert d’énergie E et du transfert de quantité de mouvement Q. Des informations dynamiques et structurelles sont obtenues. La spectroscopie neutronique mesure la même intensité intégrée S(Q) pour le transfert d’énergie (c.-à-d. le changement de vitesse des neutrons dû à la diffusion de l’échantillon, S(Q,ω) = S(Q, E), également appelé facteur de structure dynamique)9.
Pour calculer la diffusion à partir d’un matériau, il est plus approprié d’utiliser la fonction de corrélation de paire 7,10. Dans le cas de la diffraction, la fonction de corrélation de paire statique G(r) donne la probabilité de trouver le centre d’une particule à une distance donnée r du centre d’une autre particule. La spectroscopie généralise la fonction de corrélation des paires statiques et inclut l’énergie/fréquence/temps dans l’équation de diffusion. La fonction de corrélation de paire G(r) devient une fonction de temps G(r, t), qui peut être décomposée en une fonction de corrélation de paire d’atomes distincte GD(r, t) et une fonction d’auto-corrélation GS(r, t). Ceux-ci décrivent deux types de corrélations : les mouvements d’atomes corrélés par paires qui régissent la diffusion cohérente, et l’auto-corrélation qui régit la diffusion incohérente10.
La diffusion cohérente est la diffusion à partir de « la moyenne » et dépend de la phase relative des ondes diffusées. Dans le régime de diffusion à petit angle, les ondes neutroniques diffusées à partir de différents centres de diffusion (différents atomes) interfèrent de manière constructive (ont des phases similaires), et le mouvement collectif des atomes est observé avec une forte amélioration de l’intensité. La diffusion cohérente décrit essentiellement la diffusion d’un seul neutron à partir de tous les noyaux de l’échantillon10.
Lorsqu’aucune interférence constructive ne se produit entre les ondes neutroniques diffusées de différents centres, un seul atome est suivi dans le temps, et l’auto-corrélation entre la position de l’atome au temps t = 0 et le même atome au temps t est observée. Ainsi, l’information sur les positions relatives des atomes est perdue et l’accent est mis uniquement sur les fluctuations locales. La diffusion à partir des fluctuations locales régit la diffusion incohérente. La diffusion incohérente est isotrope, contribue au signal de fond et dégrade le rapport signal-bruit10,11.
En combinant tout ce qui précède, nous distinguons quatre grands processus de diffusion de neutrons10: (1) cohérent élastique (mesure les corrélations des positions atomiques), (2) cohérent inélastique (mesure les mouvements collectifs des atomes), (3) incohérent élastique (contribue à l’arrière-plan, réduit l’intensité de diffusion par le facteur de Debye-Waller (DWF) et mesure le facteur de structure incohérente élastique (EISF), décrivant la géométrie des mouvements diffusifs en géométrie confinée, et (4) incohérent inélastique (mesure la dynamique et l’auto-corrélation d’un seul atome).
Les processus dynamiques auxquels les neutrons peuvent accéder en biologie vont de l’amortissement des vibrations atomiques et moléculaires à basse fréquence, de l’interaction des molécules de solvant avec les bio-surfaces et des processus de diffusion dans la couche d’hydratation des macromolécules et de la géométrie confinée, aux mouvements diffusifs translationnels, rotationnels et tumbling à courte portée, aux domaines protéiques et aux mouvements allostériques1 . La grande diversité des méthodes et des instruments neutroniques pour mesurer la dynamique des protéines est basée sur la façon dont l’achromatisation du faisceau de neutrons incident ou sortant est réalisée et sur la façon dont l’analyse énergétique des neutrons diffusés est effectuée. Des spectromètres à triple axe aux spectromètres à temps de vol, en passant par la rétrodiffusion et l’écho de spin, on peut explorer des processus dynamiques avec des temps caractéristiques compris entre 1 x 10-14 s et 1 x 10-6 s (femtosecondes à microsecondes)12.
Le Laboratoire national d’Oak Ridge, avec ses deux sources de neutrons renommées, la source de neutrons de spallation – SNS13 et le réacteur à flux isotopique élevé – HFIR14, possède l’une des meilleures suites de spectromètres pour étudier la dynamique des biomatériaux. Parmi les exemples les plus éloquents, citons l’utilisation du spectromètre à neutrons froids (CNCS) de SNS15 pour étudier la perturbation dynamique de l’eau d’hydratation autour de la protéine fluorescente verte dans la solution16 ou les vibrations collectives sub-picosecondes de plusieurs protéines17. Un problème récurrent des recherches sur la diffusion inélastique des neutrons est que certains processus biologiques sont trop lents pour être observés. Sans configurations extrêmes qui entraînent une énorme perte d’intensité neutronique, les spectromètres à temps de vol sont limités à une résolution d’énergie de 10 μeV, correspondant à une échelle de temps maximale d’environ 200 ps10,11. Cela ne suffit pas pour observer des mouvements à grande échelle dans les protéines. Par conséquent, des instruments avec une résolution d’énergie plus élevée comme les spectromètres à rétrodiffusion sont souvent nécessaires. La combinaison des techniques de temps de vol et de rétrodiffusion s’est avérée puissante pour étudier le changement de dynamique interne du cytochrome P450cam (CYP101), une enzyme qui catalyse l’hydroxylation camphre18.
La diffusivité microscopique mesurée par le spectromètre à rétrodiffusion de SNS-BASIS19 était étonnamment bien définie et pouvait être séparée en diffusivité de l’eau (hydratation, cytoplasmique et eau de type vrac) et en diffusivité des constituants cellulaires chez les vers plats planaires, le premier animal vivant à être étudié par diffusion neutronique20 . La rétrodiffusion est une technique spectroscopique à haute résolution, mais elle est également limitée à plusieurs μeV = plusieurs nanosecondes, tandis que la dynamique lente dans les biomatériaux se manifeste également par le temps de survie de la corrélation entre la position atomique ou les orientations de spin (par exemple, les processus de relaxation, qui se produisent régulièrement dans la gamme de temps de dix à des centaines de nanosecondes).
La spectroscopie d’écho de spin neutronique (NSE) est la seule technique de diffusion neutronique à atteindre une résolution aussi élevée. Contrairement à d’autres techniques neutroniques, NSE ne nécessite pas d’achromatisation du faisceau puisqu’il utilise la phase mécanique quantique des neutrons, qui est leurs moments magnétiques. La manipulation des moments magnétiques permet l’utilisation d’une large distribution de longueur d’onde du faisceau de neutrons, tandis que la technique est sensible à de très petits changements de vitesse des neutrons de l’ordre de 1 x 10-4. NSE a été utilisé avec succès pour étudier la dynamique lente des protéines en solution pour de nombreuses protéines. Parmi ces nombreuses études pionnières, nous reconnaissons l’étude de la flexibilité segmentaire de l’immunoglobuline21 porcine; les mouvements du domaine couplé dans la Taq polymérase22; les mouvements du domaine dans le tétramère de l’alcool déshydrogénasede levure 23; le changement de conformation dans la phosphoglycérate kinase lors de la liaison du substrat3; l’activation des mouvements du domaine et la propagation dynamique des signaux allostériques dans la protéine 4,24,25 du cofacteur régulateur de l’échange Na+/H+ (NHERF1); la dynamique d’un état compact de l’ion mercurique réductase26; et la diffusion de l’hémoglobine dans les globules rouges27. Deux études plus récentes sur la dynamique des protéines ont exposé la flexibilité de l’anticorps humain Immunoglobuline G (IgG) en tant que source entropique28 et les caractéristiques de la contribution du solvant à la dynamique de la protéine de base de la myéline intrinsèquement désordonnée (MBP)5.
Le présent article explique les principes de base de la NSE, les multiples méthodes préparatoires recommandées pour une étude approfondie de la dynamique des protéines, ainsi que la méthodologie et le protocole expérimental pour l’acquisition de données NSE au spectromètre NSE du SNS, SNS-NSE. Le protocole caractérise deux protéines : l’IgG, une protéine d’anticorps humains ordinaire, et la protéine MBP intrinsèquement désordonnée. Les implications biophysiques, la pertinence des exemples pour la recherche et les limites de la technique sont brièvement discutées.
Spectroscopie NSE, la méthode pour les mesures de dynamique lente
NSE est une technique polarisée qui utilise le temps de vol des neutrons pour mesurer l’échange d’énergie (perte de polarisation) dû à l’interaction quasi-élastique entre les neutrons et les atomes dans un échantillon. Au cœur de la spectroscopie NSE se trouvent deux principes de base : (1) la capacité du spin neutronique à précéder dans le champ magnétique avec une fréquence proportionnelle à la force 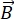 magnétique, à savoir la fréquence de Larmor29, et (b) l’écho de spin ou écho de Hann, représentant la manipulation et le recentrage du signal de polarisation lors de l’application d’une série d’impulsions de radiofréquence30.
magnétique, à savoir la fréquence de Larmor29, et (b) l’écho de spin ou écho de Hann, représentant la manipulation et le recentrage du signal de polarisation lors de l’application d’une série d’impulsions de radiofréquence30.
Les bases du processus NSE peuvent être résumées en quelques étapes simples 6,11 à l’aide de la figure 1. (1) Le faisceau de neutrons produit par la source (position 1) est polarisé (position 2), guidé et transporté (position 3), et arrive à l’entrée du spectromètre NSE, où il est tourné de 90° par la première nageoire pi-half (position 4). (2) Le faisceau polarisé (p. ex., les moments magnétiques neutroniques) devient perpendiculaire aux lignes de champ magnétique du premier aimant (première zone de précession, position 5) et commence à précession. (3) À l’extrémité de l’aimant, les spins neutroniques accumulent un certain angle de précession proportionnel à l’intensité du champ magnétique et au temps de vol passé à l’intérieur (essentiellement inversement proportionnel à la vitesse du neutron). Les vitesses individuelles des neutrons sont codées dans leur angle de précession à la fin de la première zone de précession. (4) Près de la position de l’échantillon, le pi-flipper (position 6) inverse l’orientation du spin de 180°, modifiant le signe de l’angle de précession. (5) Les neutrons interagissent avec les molécules de l’échantillon (position 7) et se dispersent. (6) Les neutrons diffusés pénètrent et précèdent dans la deuxième zone de précession (position 8) mais deviennent orientés vers l’inverse. (7) Une autre nageoire pi-half (position 9) est utilisée pour faire pivoter l’orientation du spin de perpendiculaire à la direction horizontale. Cela arrêtera la précession, traduisant l’angle de précession φ en polarisation proportionnelle au cos(φ). (8) L’analyseur (position 10) sélectionne les neutrons en fonction d’une orientation. Si l’interaction avec l’échantillon est élastique, la vitesse du neutron ne changera pas. Les neutrons passeront un temps identique à voler dans les première et deuxième zones de précession, et les angles de précession accumulés sont entièrement récupérés. La polarisation complète est restaurée sur le détecteur (position 11) en tant qu’écho de la polarisation d’origine (c’est-à-dire spin-écho). (9) Cependant, dans NSE, la diffusion est quasi-élastique, de sorte qu’un petit échange d’énergie entre les neutrons et les molécules d’échantillon conduit à des vitesses de neutrons différentes après diffusion par l’échantillon. En raison des différentes vitesses, les neutrons passeront un temps supplémentaire à voler à travers la deuxième zone de précession et n’auront pas correctement récupéré leur angle de précession. Une polarisation partielle est récupérée sur le détecteur, et la perte de polarisation due à la relaxation du spin est proportionnelle à la transformation de Cos-Fourier de la fonction spectrale S(Q, ω), la fonction de diffusion intermédiaire F(Q, t). (10) Le paramètre de temps de la fonction F(Q, t) est proportionnel à l’intensité du champ magnétique de précession. Le balayage de la perte de polarisation en fonction de l’intensité du champ magnétique donne donc une fonction de relaxation qui dépend des processus dynamiques au sein de l’échantillon.

Figure 1 : Photographie du spectromètre NSE au SNS (SNS-NSE) et schéma de trajectoire de vol des neutrons avec les composants fonctionnels les plus importants. De droite à gauche : 1 = source de neutrons ; 2 = choppers-bender-polarizer-système d’obturateur secondaire; 3 = guides de transport de faisceau; 4 = pi/2 flipper pour le premier spin-turn à 90° ; 5 = première zone de précession; 6 = pi flipper pour un spin-turn de 180°; 7 = surface de l’échantillon et environnement de l’échantillon (ici, le cryo-four est montré); 8 = deuxième zone de précession; 9 = pi/2 flipper pour le deuxième spin-turn à 90° ; 10 = analyseur; 11 = détecteur. (Notez que les parties de 3, ainsi que 2 et 1, sont situées derrière le mur bleu à l’intérieur du blindage; les hachoirs sont remplacés par un sélecteur de vitesse pour le NSE basé sur un réacteur). Veuillez cliquer ici pour voir une version agrandie de cette figure.
