Die metamere Segmentierung von Organismen ist in der Natur weit verbreitet. Wiederholte Strukturen sind essentiell für die Funktionalität von Seitenorganen wie Wirbeln, Muskeln, Nerven, Gefäßen, Gliedmaßen oder Blättern in einem Körperplan1. Als Ergebnis solcher physiologischen und geometrischen Einschränkungen der axialen Symmetrie weisen die meisten Phyla von Bilateria – wie Anneliden, Arthropoden und Chordaten – eine Segmentierung ihres embryonalen Gewebes (z. B. Ektoderm, Mesoderm) antero-posterior auf.
Wirbeltierembryonen segmentieren ihr paraxiales Mesoderm entlang der Hauptkörperachse sequenziell in Somiten mit artspezifischen Intervallen, Zählungen und Größenverteilungen. Trotz dieser Robustheit bei einzelnen Embryonen innerhalb einer Art ist die somite Segmentierung zwischen Wirbeltierarten vielseitig. Die Segmentierung erfolgt in einem riesigen Regime von Zeitintervallen (von 25 min bei Zebrafischen bis 5 h beim Menschen), Größen (von ~20 μm bei Schwanz somiten von Zebrafischen bis zu ~200 μm bei Stamm somites von Mäusen) und Anzahl (von 32 bei Zebrafischen bis ~300 bei Kornschlangen)2. Interessanterweise können sich Fischembryonen in einem weiten Temperaturbereich entwickeln (von ~ 20,5 ° C bis zu 34 ° C für Zebrafische), während sie ihre Somiten mit den richtigen Größenverteilungen intakt halten, indem sie sowohl Segmentierungsintervalle als auch axiale Dehnungsgeschwindigkeiten ausgleichen. Über diese interessanten Merkmale hinaus bleibt Zebrafisch ein nützlicher Modellorganismus, um die Segmentierung bei Wirbeltieren aufgrund der äußeren, synchronen und transparenten Entwicklung einer Fülle von Geschwisterembryonen sowie ihrer zugänglichen genetischen Werkzeuge zu untersuchen. Aus mikroskopischer Sicht entwickeln sich Teleost-Embryonen auf einem sperrigen kugelförmigen Eigelb, dehnen und runden das gastrulierende Gewebe um es herum (Abbildung 1A). In diesem Artikel stellen wir eine abgeflachte 3D-Gewebe-Explant-Kultur für Zebrafischschwänze vor. Dieses Explant-System umgeht die sphärischen Einschränkungen der Dottermasse und ermöglicht den Zugang zu hochauflösenden Live-Imagings von Fischembryonen für die somite Musterung.
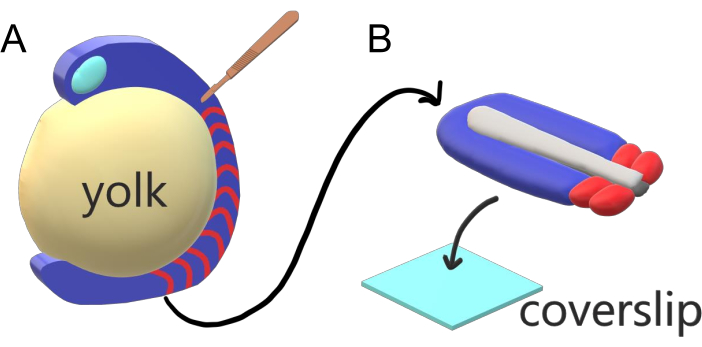
Abbildung 1: Diakammer-Explant-System für Zebrafischembryonen. (A) Zebrafischembryonen haben Vorteile für die Live-Bildgebung, wie die Transparenz von gastrulierendem embryonalem Gewebe (blau), aber das Gewebe bildet sich um eine sperrige kugelförmige Eigelbmasse (gelb), die eine nahezu objektive, hochauflösende Bildgebung in intakten Embryonen verhindert. Schwanzentstellungen können seziert werden, beginnend mit einem mikrochirurgischen Messer (braun), das aus dem Gewebe vor somites (rot) geschnitten wird und an der Grenze mit dem Eigelb posterior fortgesetzt wird. (B) Sezierte Schwanzausscheidungen können dorsoventral auf einen Abdeckungsrutsch (hellblau) gelegt werden; hält Neuralgewebe (hellgrau) oben und Notochord (dunkelgrau) unten. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
