La producción de la mayoría de los ARN mensajeros eucariotas (ARNm) implica la eliminación de intrones y la ligadura de exones en un proceso nuclear denominado empalme pre-ARNm1. Dos clases de complejos ARN-proteína (RNPs) dirigen el procesamiento del ARN pre-mensajero en ARNm maduro a través de complejos espliceosómicos. Una clase, los RNPs pre-mensajeros nacientes, se forma co-transcripcionalmente por la unión de proteínas RNP nucleares heterogéneas y otras proteínas de unión a ARN, incluyendo algunos miembros de la familia SR, produciendo complejos hnRNP2. Los RNPs nucleares pequeños ricos en uracilo de segunda clase (U snRNPs con SnRNAs U1, U2, U4, U5 y U6) se asocian con proteínas específicas de U y del núcleo3,4. Los snRNPs U interactúan de manera ordenada con regiones específicas de RNPs pre-mensajeros en una vía de remodelación dinámica a medida que los intrones se extirpan y los exones se ligan para producir mRNPs maduros5. Muchas proteínas nucleares adicionales participan en estos eventos de procesamiento6.
La galectina-1 (Gal1) y la galectina-3 (Gal3) son dos proteínas que son factores necesarios en la vía de empalme como lo demuestran los estudios de agotamiento-reconstitución7,8. La eliminación de ambas galectinas del empalme de extractos nucleares competentes (NE) suprime el ensamblaje del espliceosoma y la actividad de empalme en un paso temprano. La adición de cualquiera de las dos galectinas a un NE tan doblemente agotado restaura ambas actividades. Gal1 y Gal3 son componentes de los empaliceosomas activos, como lo demuestra la inmunoprecipitación específica de pre-ARNm, intermedios de empalme y ARNm maduro por antisuero específico para Gal1 o Gal39. Es importante destacar que Gal3 se asocia con partículas endógenas U snRNA que contienen partículas en el NE fuera de la vía de empalme, como lo demuestra la precipitación de snRNPs por anti-Gal3 antisera10. Finalmente, el silenciamiento de Gal3 en células HeLa altera los patrones de empalme de numerosos genes11.
En NE preincubado para desmontar spliceosomas preformados12, los snRNPs se encuentran en múltiples complejos sedimentando en gradientes de glicerol desde 7S hasta mayores de 60S. Aunque el fraccionamiento del gradiente de glicerol es una técnica común para el aislamiento de complejos y componentes espliceosómicos (ver referencias13,14,15 por ejemplo), hemos ampliado este método caracterizando fracciones específicas mediante inmunoprecipitaciones de anticuerpos. Un sedimento snRNP a 10S contiene solo SnRNA U1 junto con Gal3. La inmunoprecipitación de la fracción 10S con antisueros específicos para Gal3 o U1 snRNP co-precipita tanto U1 como Gal3 indicando que algunas de las monopartículas U1 snRNP están unidas a Gal310. Como U1 snRNP es el primer complejo que se une a pre-mRNP en el ensamblaje espliceosómico1,5, este paso representa un sitio de entrada potencial para Gal3 en la vía de empalme. Sobre esta base, demostramos que las monopartículas 10S Gal3-U1 snRNP unidas a perlas que contienen anti-Gal3 restauraron la actividad de empalme a un NE agotado U1 snRNP, estableciendo este complejo como un mecanismo por el cual Gal3 es reclutado en la vía espliceosomal16. Esto contrasta con los intentos de aislar los espliceosomas en etapas específicas de la reacción de empalme y catalogar los factores asociados17,18. En tales estudios, se determina la presencia de ciertos factores en algún momento, pero no el mecanismo por el cual se cargaron.
Anteriormente habíamos descrito en detalle la preparación de NE, el sustrato de empalme, el ensamblaje de la mezcla de reacción de empalme y el análisis de productos en nuestra documentación del papel de las galectinas en el empalme pre-ARNm19. Ahora describimos los procedimientos experimentales para el fraccionamiento de extractos nucleares para obtener una fracción enriquecida en el complejo Gal3 – U1 snRNP y para la inmunoselección de este último complejo para reconstituir la actividad de empalme en un extracto nuclear agotado de U1.
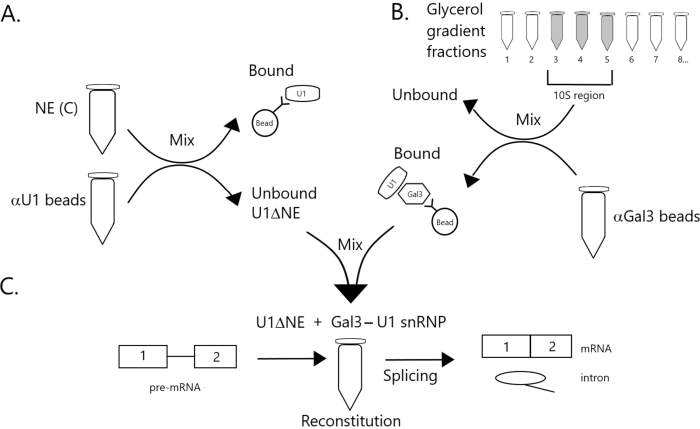
Figura 1: Diagrama esquemático que ilustra la complementación de la actividad de empalme en extracto nuclear agotado de U1 snRNP por un complejo Gal3-U1 snRNP en perlas. (A) NE en Buffer C (NE(C)) se incuba con perlas de proteína A-sefalrosa covalentemente acopladas con snRNP anti-U1 (perlas αU1). La fracción no unida se agota de U1 snRNP (U1ΔNE). (B) NE en el tampón D (NE(D)) se fracciona sobre un gradiente de glicerol de 12%-32% por ultracentrifugación. Las fracciones correspondientes a la región 10S (fracciones 3-5) se combinan y mezclan con perlas covalentemente acopladas con anticuerpos anti-Gal3 (perlas αGal3). El material unido a las perlas contiene una monopartícula Gal3-U1 snRNP. (C) El complejo snRNP Gal3-U1 de la Parte (B) se mezcla con U1ΔNE de la Parte (A) en un ensayo de empalme utilizando sustrato de pre-ARNm MINX marcado con 32P y los intermedios y productos de la reacción de empalme se analizan mediante electroforesis en gel y autorradiografía. Haga clic aquí para ver una versión más grande de esta figura.
