अधिकांश यूकेरियोटिक मैसेंजर आरएनए (एमआरएनए) के उत्पादन में एक परमाणु प्रक्रिया में इंट्रोन को हटाना और एक्सोन के बंधाव को शामिल करना शामिल है जिसे प्री-एमआरएनए स्प्लिसिंग 1 कहा जाता है। आरएनए-प्रोटीन कॉम्प्लेक्स (आरएनपी) के दो वर्ग पूर्व-मैसेंजर आरएनए के प्रसंस्करण को स्प्लिसोसोमल कॉम्प्लेक्स के माध्यम से परिपक्व एमआरएनए में निर्देशित करते हैं। एक वर्ग, नवजात पूर्व-दूत आरएनपी, विषम परमाणु आरएनपी प्रोटीन और अन्य आरएनए-बाइंडिंग प्रोटीन के बंधन द्वारा सह-ट्रांसक्रिप्शनल रूप से बनाया जाता है, जिसमें एसआर परिवार के कुछ सदस्य शामिल हैं, जो एचएनआरएनपी कॉम्प्लेक्स 2 की उपज देते हैं। दूसरी श्रेणी, यूरासिल-समृद्ध छोटे परमाणु आरएनपी (U1, U2, U4, U5, और U6 snRNAs के साथ U snRNPs) यू-विशिष्ट और कोर प्रोटीन 3,4 के साथ जुड़ा हुआ है। यू snRNPs एक गतिशील remodeling मार्ग में पूर्व मैसेंजर RNPs के विशिष्ट क्षेत्रों के साथ एक आदेशित फैशन में बातचीत के रूप में introns excised रहे हैं और exons परिपक्व mRNPs5 का उत्पादन करने के लिए ligated रहे हैं. कई अतिरिक्त परमाणु प्रोटीन इन प्रसंस्करण घटनाओं में भाग लेते हैं6।
गैलेक्टिन -1 (Gal1) और गैलेक्टिन -3 (Gal3) दो प्रोटीन हैं जो स्प्लिसिंग मार्ग में आवश्यक कारक हैं जैसा कि कमी-पुनर्गठन अध्ययन7,8 द्वारा दिखाया गया है। सक्षम परमाणु अर्क (एनई) को स्प्लिसिंग से दोनों गैलेक्टिन को हटाने से स्प्लिसोसोम असेंबली और स्प्लिसिंग गतिविधि को शुरुआती चरण में समाप्त कर दिया जाता है। इस तरह के दोगुने समाप्त एनई में या तो गैलेक्टिन के अलावा दोनों गतिविधियों को बहाल करता है। Gal1 और Gal3 सक्रिय spliceosomes के घटक हैं जैसा कि पूर्व-mRNA के विशिष्ट immunoprecipitation, splicing intermediates, और परिपक्व mRNA द्वारा या तो Gal1 या Gal39 के लिए विशिष्ट एंटीसीरम द्वारा प्रमाणित है। महत्वपूर्ण रूप से, Gal3 अंतर्जात U snRNA के साथ सहयोग करता है जिसमें splicing pathway के बाहर NE में कण होते हैं, जैसा कि एंटी-Gal3 antisera10 द्वारा snRNPs की वर्षा द्वारा दिखाया गया है। अंत में, हेला कोशिकाओं में Gal3 की चुप्पी कई जीन11 के splicing पैटर्न को बदल देती है।
पूर्वनिर्मित spliceosomes12 को अलग करने के लिए एनई प्री-इनक्यूबेटेड में, snRNPs कई परिसरों में पाए जाते हैं जो 7S से 60S से अधिक तक ग्लिसरॉल ग्रेडिएंट में तलछट करते हैं। यद्यपि ग्लिसरॉल ग्रेडिएंट फ्रैक्शनेशन स्प्लिसोसोमल कॉम्प्लेक्स और घटकों के अलगाव के लिए एक आम तकनीक है (उदाहरण के लिए संदर्भ 13,14,15 देखें), हमने एंटीबॉडी इम्यूनोप्रिसिपेशन का उपयोग करके विशिष्ट अंशों की विशेषता करके इस विधि का विस्तार किया है। 10S पर एक snRNP तलछट में Gal3 के साथ केवल U1 snRNA होता है। Gal3 या U1 snRNP सह-अवक्षेप दोनों U1 और Gal3 के लिए विशिष्ट antisera के साथ 10S अंश के immunoprecipitation U1 और Gal3 दोनों को दर्शाता है कि U1 snRNP मोनोपार्टिकल्स में से कुछ Gal310 के लिए बाध्य हैं। U1 snRNP के रूप में पहला जटिल है कि spliceosomal assembly1,5 में पूर्व-mRNP के लिए बांधता है, यह कदम splicing मार्ग में Gal3 के लिए एक संभावित प्रवेश साइट का प्रतिनिधित्व करता है। इस आधार पर, हमने दिखाया कि 10S Gal3-U1 snRNP मोनोपार्टिकल्स एंटी-गैल 3 से बंधे हैं, जिसमें मोतियों ने U1 snRNP को समाप्त करने के लिए splicing गतिविधि को बहाल किया है, इस परिसर को एक तंत्र के रूप में स्थापित किया गया है जिसके द्वारा Gal3 को spliceosomal pathway16 में भर्ती किया जाता है। यह splicing प्रतिक्रिया में विशिष्ट चरणों में spliceosomes को अलग करने और संबंधित कारकों को सूचीबद्ध करने के प्रयासों के साथ विरोधाभास करता है17,18। इस तरह के अध्ययनों में, कुछ समय बिंदु पर कुछ कारकों की उपस्थिति का पता लगाया जाता है लेकिन उस तंत्र को नहीं जिसके द्वारा उन्हें लोड किया गया था।
हमने पहले एनई की तैयारी, स्प्लिसिंग सब्सट्रेट, स्प्लिसिंग प्रतिक्रिया मिश्रण की असेंबली, और प्री-एमआरएनए स्प्लिसिंग 19 में गैलेक्टिन की भूमिका के हमारे प्रलेखन में उत्पादों के विश्लेषण का विस्तार से वर्णन किया था। अब हम परमाणु अर्क के फ्रैक्शनेशन के लिए प्रयोगात्मक प्रक्रियाओं का वर्णन करने के लिए Gal3 – U1 snRNP परिसर में समृद्ध एक अंश प्राप्त करने के लिए और बाद के परिसर के immuno-चयन के लिए एक U1-depleted परमाणु निकालने में splicing गतिविधि का पुनर्गठन करने के लिए।
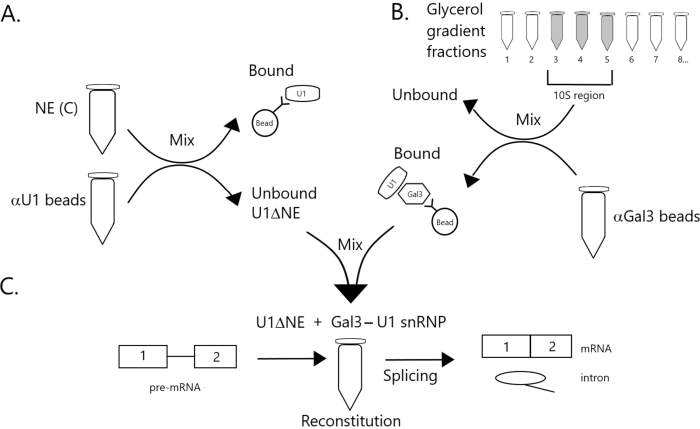
चित्र 1: परमाणु अर्क में स्प्लिसिंग गतिविधि के पूरक को दर्शाने वाला योजनाबद्ध आरेख मोती पर एक Gal3-U1 snRNP परिसर द्वारा U1 snRNP के समाप्त हो गए हैं। (A) बफर C (NE(C)) में NE को प्रोटीन A-Sepharose मोतियों के साथ सहसंयोजक रूप से एंटी-U1 snRNP (αU1 मोतियों) के साथ युग्मित किया गया है। अनबाउंड भिन्न U1 snRNP (U1ΠNE) से समाप्त हो गया है। (बी) बफर डी (एनई (डी)) में एनई को अल्ट्रासेंट्रीफ्यूजेशन द्वारा 12% -32% ग्लिसरॉल ग्रेडिएंट पर विभाजित किया जाता है। 10S क्षेत्र (अंश 3-5) के अनुरूप अंशों को संयुक्त किया जाता है और मोतियों के साथ मिश्रित किया जाता है जो एंटी-गैल 3 एंटीबॉडी (αGal3 मोतियों) के साथ सहसंयोजक रूप से युग्मित होते हैं। मोतियों के लिए बाध्य सामग्री एक Gal3-U1 snRNP मोनोपार्टिकल शामिल हैं। (ग) भाग (बी) से Gal3-U1 snRNP परिसर को भाग (ए) से U1ΠNE के साथ मिश्रित किया जाता है, जिसमें 32P-लेबल वाले MINX पूर्व-mRNA सब्सट्रेट का उपयोग किया जाता है और splicing प्रतिक्रिया के मध्यवर्ती और उत्पादों का विश्लेषण जेल वैद्युतकणसंचलन और ऑटोरेडियोग्राफी द्वारा किया जाता है। कृपया इस आंकड़े का एक बड़ा संस्करण देखने के लिए यहाँ क्लिक करें.
