Производство большинства эукариотических мессенджерных РНК (мРНК) включает удаление интронов и лигирование экзонов в ядерном процессе, называемом сплайсингом премрнК1. Два класса РНК-белковых комплексов (RNP) направляют обработку пре-мессенджерной РНК в зрелую мРНК через сплайсеосомальные комплексы. Один класс, зарождающиеся предмаршированные RNP, образуется котранскрипционно путем связывания гетерогенных ядерных белков RNP и других РНК-связывающих белков, включая некоторых членов семейства SR, давая комплексы hnRNP2. Второй класс, богатый урацилом малые ядерные RNP (U snRNP с U1, U2, U4, U5 и U6 snRNAs) связан с U-специфическими и основными белками3,4. U snRNP взаимодействуют упорядоченным образом с определенными областями предмарш-мессенджерных RNP в динамическом пути ремоделирования, поскольку интроны иссекаются, а экзоны лигируются для получения зрелых mRNPs5. Многие дополнительные ядерные белки участвуют в этих событиях обработки6.
Галектин-1 (Gal1) и галектин-3 (Gal3) являются двумя белками, которые являются необходимыми факторами в пути сплайсинга, как показано исследованиями истощения-восстановления7,8. Удаление обоих галектинов из сращивания компетентных ядерных экстрактов (NE) отменяет сплайсеосомную сборку и сплайсинговую деятельность на ранней стадии. Добавление любого галектина к такому вдвойне истощенному NE восстанавливает обе активности. Gal1 и Gal3 являются компонентами активных сплайсеосом, о чем свидетельствует специфическая иммунопреципитация пре-мРНК, сплайсинговых промежуточных продуктов и зрелой мРНК антисывороткой, специфичной для Gal1 или Gal39. Важно отметить, что Gal3 ассоциируется с эндогенной U snRNA, содержащей частицы в NE вне пути сплайсинга, как показано осаждением snRNP анти-Gal3 antisera10. Наконец, глушение Gal3 в клетках HeLa изменяет паттерны сплайсинга многочисленных генов11.
В NE, предварительно инкубированном для разборки предварительно сформированных сплайсеосом12, snRNP обнаруживаются в нескольких комплексах, осажденных в градиентах глицерина от 7S до более 60S. Хотя фракционирование градиента глицерина является распространенным методом выделения сплайсеосомальных комплексов и компонентов (см., например, ссылки13,14,15), мы расширили этот метод, охарактеризовав специфические фракции с использованием иммунопреципитаций антител. SnRNP осадок при 10S содержит только U1 snRNA вместе с Gal3. Иммунопреципитация фракции 10S с антисыворотки, специфичной для Gal3 или U1 snRNP, осаждает как U1, так и Gal3, указывая на то, что некоторые из моночастиц U1 snRNP связаны с Gal310. Поскольку U1 snRNP является первым комплексом, который связывается с pre-mRNP в сплайсеосомальной сборке1,5, этот шаг представляет собой потенциальный входной участок для Gal3 в путь сплайсинга. На этой основе мы показали, что моночастицы 10S Gal3-U1 snRNP, связанные с бусинами, содержащими анти-Gal3, восстанавливают активность сплайсинга к U1 snRNP истощенной NE, устанавливая этот комплекс как один механизм, с помощью которого Gal3 набирается в сплайсеосомальный путь16. Это контрастирует с попытками выделения сплайсеосом на определенных этапах реакции сплайсинга и каталогизации связанных с ними факторов17,18. В таких исследованиях констатируется наличие определенных факторов в какой-то момент времени, но не механизм, с помощью которого они были загружены.
Ранее мы подробно описали подготовку NE, сплайсинговую подложку, сборку реакционной смеси сплайсинга и анализ продуктов в нашей документации о роли галектинов в сплайсинге премрнК19. Теперь описаны экспериментальные процедуры фракционирования ядерных экстрактов для получения фракции, обогащенной комплексом SnRNP Gal3 – U1, и иммуноотбора последнего комплекса для восстановления сплайсинговой активности в ядерном экстракте, обедненном U1.
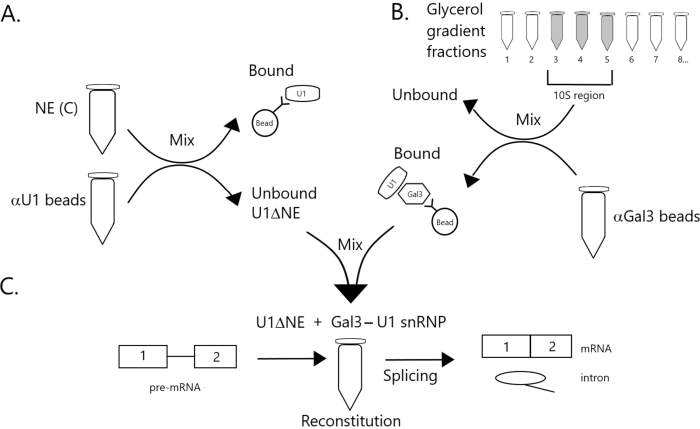
Рисунок 1: Принципиальная диаграмма, иллюстрирующая дополнение сплайсинговой активности в ядерном экстракте, обедненном U1 snRNP комплексом Gal3-U1 snRNP на шариках. (A) NE в буфере C (NE(C)) инкубируют с белком A-Sepharose бусинами, ковалентно связанными с анти-U1 snRNP (бусины αU1). Несвязанная фракция обедняется U1 snRNP (U1ΔNE). (B) NE в буфере D (NE(D)) фракционируют по 12%-32% градиенту глицерина путем ультрацентрифугирования. Фракции, соответствующие области 10S (фракции 3-5), объединяют и смешивают с бусинами, ковалентно связанными с антителами против Gal3 (шарики αGal3). Материал, связанный с шариками, содержит моночастицу Gal3-U1 snRNP. (C) Комплекс SnRNP Gal3-U1 из части (B) смешивают с U1ΔNE из части (A) в сплайсинговом анализе с использованием 32P-меченого субстрата MINX pre-mRNA, а промежуточные продукты и продукты реакции сплайсинга анализируют с помощью гелевого электрофореза и ауторадиографии. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
