Produktion av mest eukaryotic budbärareRNA (mRNAs) innebär borttagning av introner och ligation av exons i en kärn- processaa som benämns pre-mRNA-skarvning1. Två klasser av RNA-proteinkomplex (RNPs) styr bearbetningen av pre-messenger RNA till mogna mRNA via spliceosomal komplex. En klass, gryende pre-messenger RNPs, bildas co-transcriptionally genom bindning av heterogena nukleära RNP-proteiner och andra RNA-bindande proteiner, inklusive vissa medlemmar av SR-familjen, vilket ger hnRNP-komplex2. Den andra klassen, uracil-rika små nukleära RNPs (U snRNPs med U1, U2, U4, U5 och U6 snRNAs) är associerad med U-specifika och kärnproteiner3,4. U snRNPs interagerar på ett ordnat sätt med specifika regioner av pre-messenger RNPs i en dynamisk ombyggnadsväg som introner är strukna och exons är ligated för att producera mogna MRNPs5. Många ytterligare nukleära proteiner deltar i dessa bearbetningsevenemang6.
Galectin-1 (Gal1) och galectin-3 (Gal3) är två proteiner som krävs faktorer i skarvningsvägen som framgår av utarmnings-rekonstitutionsstudier7,8. Avlägsnande av båda galectins från skarvning kompetent kärnämne extrakt (NE) avskaffar skareosome montering och splitsning verksamhet i ett tidigt steg. Tillägg av antingen galectin till ett så dubbelt utarmat NE återställer båda aktiviteterna. Gal1 och Gal3 är komponenter i aktiva splitseomer, vilket framgår av specifik immunprecipitation av pre-mRNA, skarvning av intermediärer och mogna mRNA med antiserumspecifikt för antingen Gal1 eller Gal39. Viktigt är att Gal3 associerar med endogent U snRNA som innehåller partiklar i NE utanför skarvningsvägen som framgår av utfällning av snRNPs av anti-Gal3 antisera10. Slutligen, ljuddämpning av Gal3 i HeLa celler förändrar skarvning mönster av många gener11.
I NE pre-incubated för att demontera förformade spliteosomer12, snRNPs finns i flera komplex sedimentering i glycerol gradienter från 7S till mer än 60S. Även om glycerol gradient fraktionering är en vanlig teknik för isolering av spliceosomal komplex och komponenter (se referenser13,14,15 till exempel), har vi utvidgat denna metod genom att karakterisera specifika fraktioner med hjälp av antikroppar immunprecipitations. En snRNP sedimentering vid 10S innehåller endast U1 snRNA tillsammans med Gal3. Immunoprecipitation av 10S-fraktionen med antisera specifik för Gal3 eller U1 snRNP co-fäller ut både U1 och Gal3 som indikerar att några av U1 snRNP-monopartiklarna är bundna till Gal310. Eftersom U1 snRNP är det första komplexet som binder till pre-mRNP i spliteosomal sammansättning1,5, representerar detta steg en potentiell ingångsplats för Gal3 i skarvningsvägen. På denna grund visade vi att 10S Gal3-U1 snRNP monopartiklar bundna till anti-Gal3 som innehåller pärlor återställde splitsning aktivitet till en U1 snRNP utarmade NE, upprätta detta komplex som en mekanism genom vilken Gal3 rekryteras till den spliteosomala vägen16. Detta står i kontrast till försök att isolera splitsosomer i specifika stadier i skarvningsreaktionen och katalogisering av de associerade faktorerna17,18. I sådana studier fastställs förekomsten av vissa faktorer vid någon tidpunkt men inte den mekanism genom vilken de laddades.
Vi hade tidigare beskrivit i detalj beredningen av NE, skarvning substratet, monteringen av skarvning reaktion blandning och analys av produkter i vår dokumentation av galectins roll i pre-mRNA splitsning19. Vi beskriver nu de experimentella förfarandena för fraktionering av kärnextrakt för att erhålla en bråkdel berikad i Gal3 – U1 snRNP komplex och för immuno-urval av det senare komplexet att rekonstituerande splitsning verksamhet i en U1-utarmade kärnämne extrakt.
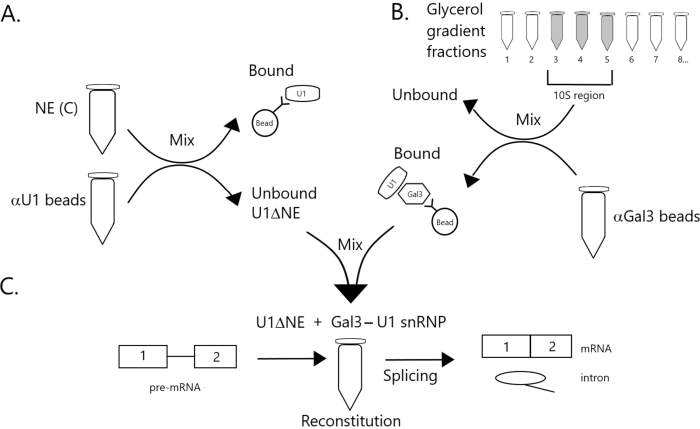
Figur 1: Schematiskt diagram som illustrerar komplementering av skarvningsaktivitet i kärnextrakt som uttömts av U1 snRNP av ett Gal3-U1 snRNP-komplex på pärlor. Den obundna fraktionen är uttömd av U1 snRNP (U1ΔNE). (B) NE i buffert D (NE(D)) är fraktionerad över en 12-32% glycerol gradient genom ultracentrifugation. Fraktioner som motsvarar 10S-regionen (fraktioner 3-5) kombineras och blandas med pärlor kovalent i kombination med anti-Gal3-antikroppar (αGal3 pärlor). Materialet som är bundet till pärlorna innehåller en Gal3-U1 snRNP monopartikel. C) Gal3-U1 snRNP-komplexet från del B blandas med U1ΔNE från del A i en skarvningsanalys med 32P-märkt MINX pre-mRNA-substrat och intermediärerna och produkterna i skarvningsreaktionen analyseras med gelelektrofores och autoradiografi. Klicka här för att se en större version av den här figuren.
