La señalización reactiva de especies de oxígeno (ROS) regula el desarrollo y el funcionamiento del sistema nervioso1. Una fuente celular importante de ROS son las NADPH oxidasas (NOX), que son proteínas transmembrana que generan superóxido y peróxido de hidrógeno(H2O2)2. Las enzimas NOX se encuentran en todo el sistema nervioso central (SNC), y el ROS derivado de NOX contribuye al desarrollo neuronal3,4,5,6. Se ha demostrado que el mantenimiento y la diferenciación de las células madre neurales, el establecimiento de la polaridad neuronal, la excrecencia de la neurita y la plasticidad sináptica requieren niveles adecuados de ROS7,8,9,10,11. Por otro lado, la producción incontrolada de ROS por los NOXes contribuye a trastornos neurodegenerativos como la enfermedad de Alzheimer, la esclerosis múltiple y la lesión cerebral traumática12,13,14. Por lo tanto, la producción de ROS fisiológicamente relevante es fundamental para mantener condiciones saludables.
El desarrollo de biosensores codificados genéticamente facilitó enormemente la detección de ROS celular. Una ventaja importante de los biosensores codificados genéticamente es el aumento de la resolución temporal y espacial de la señal ROS, ya que estos sensores se pueden dirigir específicamente a ubicaciones distintas. GFP sensible a redox (roGFP) es un tipo de estos biosensores ROS. La variante roGFP2-Orp1 detecta específicamenteH2O2 a través de su dominio Orp1, que es una proteína de la familia glutatión peroxiredoxina de la levadura15,16. La oxidación de la proteína Orp1 se transfiere a roGFP2 para alterar su conformación (Figura 1A). La sonda exhibe dos picos de excitación cercanos a 405 nm y 480 nm, y un solo pico de emisión a 515 nm. Tras la oxidación, la intensidad de fluorescencia alrededor de los picos de excitación cambia: mientras que la excitación de 405 nm aumenta, la excitación de 480 nm disminuye. Por lo tanto, roGFP2-Orp1 es un biosensor ratiométrico, y los niveles deH2O2se detectan por la relación de intensidades de fluorescencia excitadas en dos longitudes de onda diferentes (Figura 1B). En general, roGFP2-Orp1 es una herramienta versátil para imágenes ROS que se puede utilizar de manera eficiente in vivo.
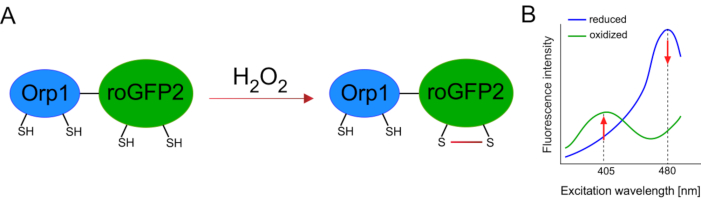
Figura 1:Representación esquemática y espectros de excitación de roGFP2-Orp1. (A)La transferencia de oxidantes se produce entre Orp1 y roGFP2 en respuesta aH2O2,dando lugar a cambios conformacionales en roGFP2. (B) Los espectros de excitación del roGFP2-Orp1 exhiben dos picos de excitación a 405 nm y 480 nm y un pico de emisión única a 515 nm. Tras la oxidación porH2O2, la excitación de 405 nm aumenta mientras que la excitación de 480 nm disminuye. Esto da lugar a una lectura ratiometric para la presencia deH2O2. La cifra ha sido modificada de Bilan y Belousov (2017)16 y Morgan et al. (2011)25. Haga clic aquí para ver una versión más amplia de esta figura.
El sistema modelo Danio rerio (pez cebra) tiene varias ventajas para aplicar biosensores codificados genéticamente. La transparencia óptica de los embriones y larvas permite la obtención de imágenes in vivo no invasivas. Se están desarrollando nuevas herramientas de imagen para lograr una mayor resolución y una penetración más profunda17. Además, existen herramientas establecidas para la manipulación genética (expresión ectópica de ARNm, transgénesis de Tol2, etc.) y la edición del genoma (TALENs, CRISPR/Cas9, etc.), que promueve la generación de animales transgénicos18. A medida que los embriones de pez cebra se desarrollan fuera de la madre, este sistema permite aún más el acceso y la manipulación de los embriones. Por ejemplo, las inyecciones en etapa unicelular y los tratamientos farmacológicos se pueden hacer fácilmente.
Aquí, utilizamos pez cebra para expresar transitoriamente el H2O2-biosensores específicos roGFP2-Orp1 mediante la inyección de ARNm transcrito in vitro. Estos embriones se pueden utilizar tanto para la obtención de imágenes in vitro de neuronas cultivadas como para la obtención de imágenes in vivo (Figura 2). Se describe un protocolo para la disección y chapado de células ganglionares de la retina (RGCs) de embriones de pez cebra seguido de la evaluación de H2O2– niveles en neuronas cultivadas. Entonces, presentamos un método para la proyección de imagen in vivo de roGFP2-Orp1-expresando embriones y larvas usando microscopia confocal. Este enfoque no sólo permite determinar los niveles fisiológicos deH2O2-sino también los cambios potenciales que ocurren en diferentes etapas o condiciones de desarrollo. En general, este sistema proporciona un método confiable para detectarH2O2 en células vivas y animales para estudiar el papel deH2O2 en el desarrollo, la salud y la enfermedad.
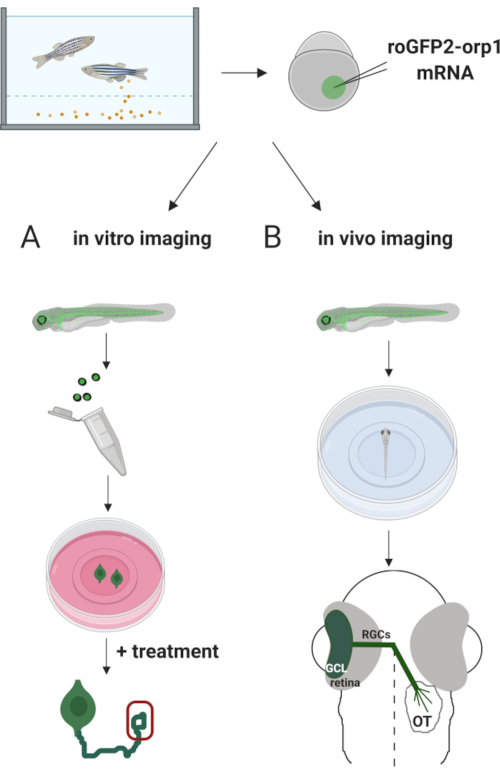
Figura 2. Esquema del enfoque experimental. Brevemente, después de la recolección de embriones, roGFP2-Orp1 ARNm se inyecta en la yema de los embriones de pez cebra en etapa unicelular. Los embriones en desarrollo se pueden utilizar tanto para(A) imágenes in vitro como(B) in vivo. (A)Los embriones GFP-positivos se utilizan para diseccionar retinas para la colección de RGC en 34 hpf. Los RGCs disociados se platean en cubrebocas recubiertas de PDL/laminina en medios ZFCM (+). La proyección de imagen del cono del crecimiento se puede conducir mientras que RGCs extiende sus axones después de 6-24 h de la galjanoplastia. Las células pueden ser sometidas a diferentes tratamientos para medir los posibles cambios en los niveles deH2O2.- Aquí, medimos H2O2-niveles en los conos de crecimiento de RGCs (rojo). (B)Los embriones positivos de GFP se utilizan para la obtención de imágenes in vivo. A la edad deseada, los embriones pueden anestesiarse y montarse en platos con fondo de vidrio de 35 mm para obtener imágenes confocales. Aquí, los embriones se montan ventralmente para la proyección de imagen retiniana. Schematic muestra el desarrollo de la retina en el pez cebra. Los RGCs forman la capa de la célula del ganglio (GCL), que es la capa más interna en retina. Los axones de RGC se convierten en nervio óptico para cruzar midline, formando quiasma óptico. Entonces, los axones de RGC crecen dorsalmente para hacer sinapsis en el tectum óptico en el mesencéfano. Haga clic aquí para ver una versión más amplia de esta figura.
