wAlbB-SG is a localized (Singapore) Ae. aegypti line stably infected with the wAlbB strain of Wolbachia. Using the protocol described in this paper, we evaluated the male mating competitiveness of an inbred and an outcrossed line of wAlbB-SG to determine if inbreeding results in a loss in male mating fitness. The inbred line had been maintained for 11 generations in the insectary, while the outcrossed line was generated by backcrossing the females with wild-type male Ae. aegypti. Males from the inbred and outcrossed lines were competed against each other for mating with wild-type wild-type female Ae. aegypti. The mating competitiveness assay was conducted in triplicate.
The results indicated that RhB did not affect the fitness of the males as the data for female insemination was not biased towards or against mating with RhB-sucrose-fed males (Table 1 and Figure 6) As RhB does not affect the mating fitness of the males, we proceed to analyse the data based on the percentage of inseminated females mated by either the inbred or outcrossed line (Table 2 and Figure 7). The result across the experimental triplicates were consistent; there was a significantly higher percentage of females mated with the outcross males than with the inbred males in all three replicates (P ≤ 0.05, Mann-Whitney U-test). These results suggest a potential loss in male mating fitness after several generations of inbreeding in the laboratory.

Figure 1: Lateral view of male (left) and female (right) Aedes aegypti pupae. Under the same rearing conditions, Ae. aegypti can be sexed at the pupal stage according to size; males are significantly smaller than females. Please click here to view a larger version of this figure.
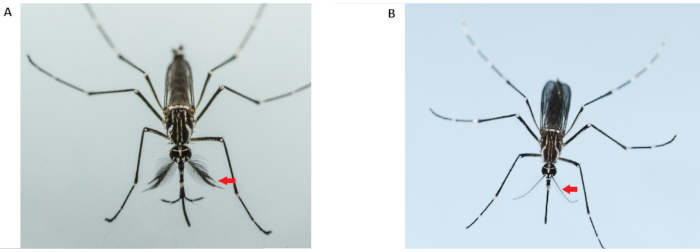
Figure 2: Differentiation of male (left) and female (right) Aedes aegypti adults. Adult male mosquitoes (left) have bushier and hairier antennae than the adult female; the red arrows indicate the antennae. Please click here to view a larger version of this figure.
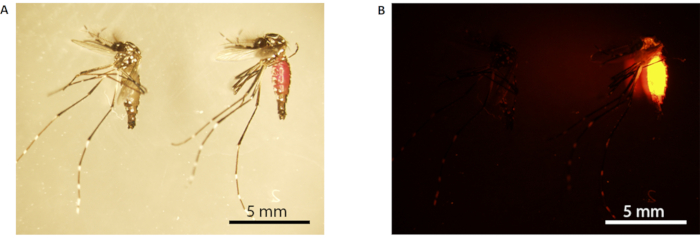
Figure 3: Rhodamine B marking of male mosquito. (A) Light microscopy; (B) fluorescence microscopy. The mosquito on the left is unmarked (fed with 10% w/v sucrose), while the one on the right is marked (fed with 0.2% RhB-sucrose). Marked mosquitoes have a visible pink abdomen under white light (the mosquito on the right in A), which fluoresces bright orange under fluorescence microscopy (B). Scale bars = 5 mm. Abbreviation: RhB = Rhodamine B. Please click here to view a larger version of this figure.
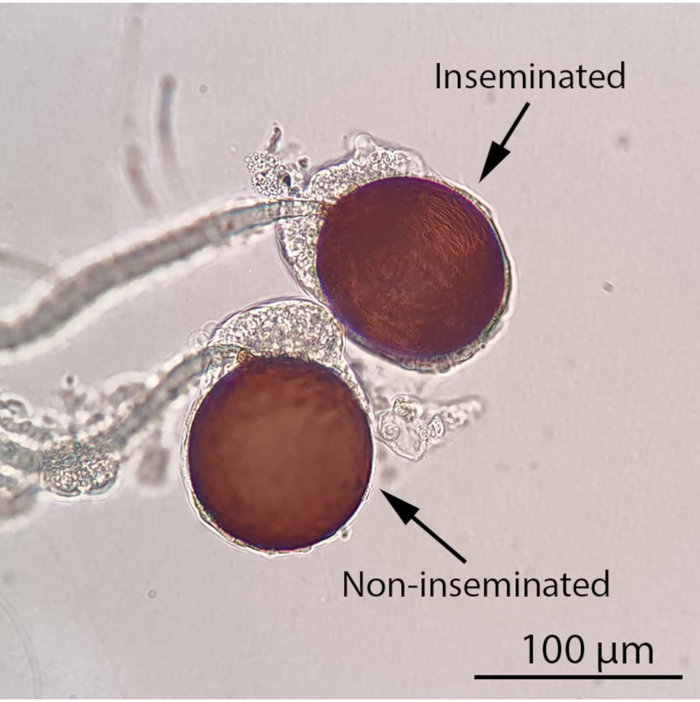
Figure 4: Inseminated and non-inseminated female spermathecae under a compound light microscope (100x magnification). The insemination status of a female mosquito can be determined by observing its spermathecae under a compound light microscope. An inseminated female mosquito will contain at least one filled spermatheca while all three spermathecae of a non-inseminated female mosquito will be empty. Thread-like, motile sperm will be visible in a filled spermatheca under a compound light microscope. Scale bar = 100 µm. Please click here to view a larger version of this figure.

Figure 5: Female mosquito spermathecae inseminated with seminal fluids under a fluorescence stereo microscope. (A) RhB-marked and (B) unmarked spermathecae inseminated with RhB-marked seminal fluids will fluoresce bright orange under fluorescence microscopy. Scale bars = 100 µm. Please click here to view a larger version of this figure.
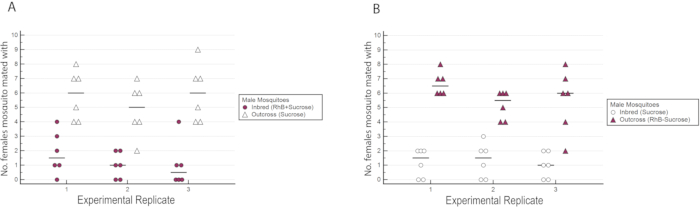
Figure 6: Number of wild-type females inseminated by either the inbred or outcross males in the experimental triplicates with reciprocal marking. (A) The inbred males were marked with RhB while the outcross males were unmarked. (B) The outcross males were marked with RhB while the inbred males were unmarked. A higher number of females was observed to have mated with outcross males regardless of their marking status. Please click here to view a larger version of this figure.
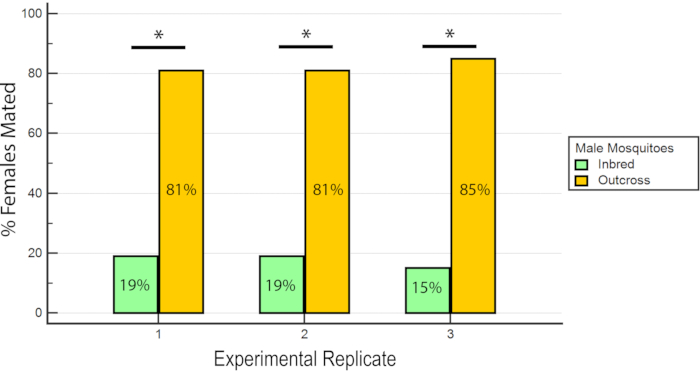
Figure 7: Proportion of inseminated females mated with inbred or outcross males in the 3 experimental replicates. For each experimental replicate, there is a significantly higher percentage of females mated with the outcross males (P ≤ 0.05, Mann-Whitney U-test). Please click here to view a larger version of this figure.
Video 1: Dissection of female Aedes aegypti for spermathecae under a light stereo microscope. Please click here to download this Video.
| ♀ x Inbred (RhB) ♂ a | ♀ x Outcross (unmarked) ♂ b | ♀ x Inbred (unmarked) ♂ c | ♀ x Outcross (RhB) ♂ d | Overall insemination rate (a+b+c+d/120) |
|
| Replicate 1 | 11 | 35 | 7 | 40 | 77.5% (93/120) |
| Replicate 2 | 6 | 29 | 8 | 31 | 61.7% (74/120) |
| Replicate 3 | 6 | 36 | 6 | 33 | 67.5% (81/120) |
Table 1: Number of females mated with RhB-marked and unmarked wAlbB-Sg Aedes aegypti inbred and outcross males. A total number of 120 females were used in each replicate.
| Percent inseminated females | ||
| Inbred males | Outcross males | |
| Replicate 1 | 19% (18/93) | 81% (75/93) |
| Replicate 2 | 19% (14/74) | 81% (60/74) |
| Replicate 3 | 15% (12/81) | 85% (69/81) |
Table 2: Percentage of inseminated females mated with wAlbB-Sg Aedes aegypti inbred and outcross males.
Supplemental Figure S1: Comparison of workflow for RhB-based and conventional mating competitiveness assay. In comparison to the conventional mating competitiveness assay, the simplified and shortened workflow for RhB-based mating competitiveness assay significantly reduces the experimental duration. Please click here to download this File.
Supplemental Figure S2: Kaplan Meier survival curves of male adult Aedes aegypti during and after feeding with 0.2% and 0.4% rhodamine B–sucrose feeding. Percent survival of (A) male wild-type and (B) wAlbB-Sg Ae. aegypti during and after three days of feeding on 0.2% and 0.4% RhB-sucrose, compared to controls that were fed with sucrose only. Please click here to download this File.
Supplemental Table S1: Insemination rate of females in a W 60 cm x D 60 cm x H 60 cm cage (ratio of 10 females to 20 males) at 1-, 2-, and 3-h time points. Please click here to download this Table.
