Moleculaire chaperones dragen bij aan eiwithomeostase, inclusief eiwitvouwing, transport en afbraak. Ze reguleren verschillende cellulaire processen en zijn gekoppeld aan tal van ziekten zoals kanker en neurodegeneratieve ziekten1. Heat shock protein 90 (Hsp90) is een van de belangrijkste chaperones waarvan de functie afhankelijk is van conformatiesveranderingen aangedreven door ATP-hydrolyse en binding met cliënteiwitten gemedieerd door zijn co-chaperones2. Ondanks een duidelijk potentieel van Hsp90 als het therapeutische doelwit, vormt het verfijnen van de functie ervan een grote uitdaging. Er zijn verschillende Hsp90-remmers gericht op het N-terminale ATP-bindingsgebied, die zijn geëvalueerd in klinische onderzoeken, maar geen van hen is goedgekeurd voor het op de markt brengen3. Vanwege het ontbreken van een goed gedefinieerde ligandbindende pocket4,heeft het richten op het C-terminale gebied van Hsp90 beperkt succes gehad4. Onlangs is verstoring van Hsp90-cochaperoninteracties door kleine moleculen onderzocht als een alternatieve strategie5. Het richten op de Hsp90-cochaperoninteracties zou geen algemene celstressrespons uitlokken en biedt de mogelijkheid om specifiek verschillende intracellulaire processen te reguleren. Het geconserveerde MEEVD pentapeptide aan de C-terminus van Hsp90 is verantwoordelijk voor de interactie met het tetratricopeptide repeat (TPR) motief van co-chaperones6. Van de 736 TPR-motiefbevattende eiwitten die zijn geannoteerd in de menselijke eiwitdatabase, interageren ~ 20 verschillende eiwitten met Hsp90 via dit peptide7. Moleculen die concurreren om MEEVD-peptidebinding zouden de interacties tussen Hsp90 en co-chaperones die een TPR-domein bevatten, verstoren. De peptidebindingsplaats heeft een vergelijkbare tertiaire structuur, maar de algehele homologie tussen verschillende TPR-motiefdomeinen is relatief laag7, wat een mogelijkheid biedt om moleculen te identificeren die specifiek in staat zijn om interacties tussen Hsp90 en bepaalde TPR-motief co-chaperones te blokkeren. Onder deze TPR-motief co-chaperones, FK506-bindend eiwit (FKBP) 51 en FKBP52 zijn regulatoren van steroïde hormoonreceptor (SHR) signalering en betrokken bij verschillende steroïde hormoon-afhankelijke ziekten, waaronder kanker, stress-gerelateerde ziekten, metabole ziekten, en de ziekte van Alzheimer8. Hoewel FKBP51 en FKBP52 > 80% sequentie-gelijkenis delen, verschillen hun functies: FKBP52 is een positieve regulator van SHR-activiteit, terwijl FKBP51 in de meeste gevallen een negatieve regulator is8. Daarom biedt het identificeren van moleculen, die specifiek interacties tussen Hsp90 en FKBP51 of FKBP52 blokkeren, een veelbelovend therapeutisch potentieel voor gerelateerde ziekten.
Eenmplified Luminescent Proximity Homogenous Assay (AlphaScreen) werd voor het eerst ontwikkeld in 1994 door Ullman EF et al.9. Nu wordt het veel gebruikt om verschillende soorten biologische interacties te detecteren, zoals peptide10,eiwit11,DNA12,RNA13en suiker14. Bij deze techniek zijn er twee soorten kralen (diameter 200 nm), de ene is de donorkraal en de andere is de acceptorkraal. De biomoleculen worden op deze kralen geïmmobiliseerd; hun biologische interacties brengen donor- en acceptorkralen in de buurt. Bij 680 nm licht een foto-ensitizer in de donorkraal op en zet zuurstof om in singletzuurstof. Omdat de singletzuurstof een korte levensduur heeft, kan deze slechts tot 200 nm diffunderen. Als de acceptorkraal in de buurt is, reageert het thioxeenderivaat met de singletzuurstof die chemiluminescentie genereert bij 370 nm. Deze energie activeert verder fluoroforen in dezelfde acceptorkraal om licht uit te stralen bij 520-620 nm15. Als de biologische interacties worden verstoord, kunnen de acceptorkraal en donorkraal de nabijheid niet bereiken, wat resulteert in het singlet zuurstofverval en een laag geproduceerd signaal.
Hier beschrijven we een protocol met behulp van deze techniek voor het screenen van kleine moleculen die interacties tussen Hsp90 en TPR co-chaperones remmen, met name FKBP51 en FKBP52. De 10 aminozuur lange peptiden die overeenkomen met Hsp90 extreme C-terminus zijn bevestigd aan acceptor kralen. Gezuiverde GST-gelabelde TPR co-chaperones interageren met glutathion-gekoppelde donorkralen. Wanneer de interactie tussen Hsp90-afgeleide peptiden en TPR-motief co-chaperones de kralen samenbrengt, wordt een versterkt signaal geproduceerd (Figuur 1A). Als de gescreende kleine moleculen de interacties tussen Hsp90 en TPR-motief co-chaperones kunnen remmen, zal dit versterkte signaal worden verminderd(figuur 1B). Hun IC50 kan worden berekend door kwantitatieve meting. Dit protocol kan worden uitgebreid naar alle chaperonne – TPR-motief co-chaperonne interacties van belang en is van groot belang bij de ontwikkeling van nieuwe moleculen, met name het blokkeren van de interactie tussen Hsp90 en FKBP51 of FKBP52.
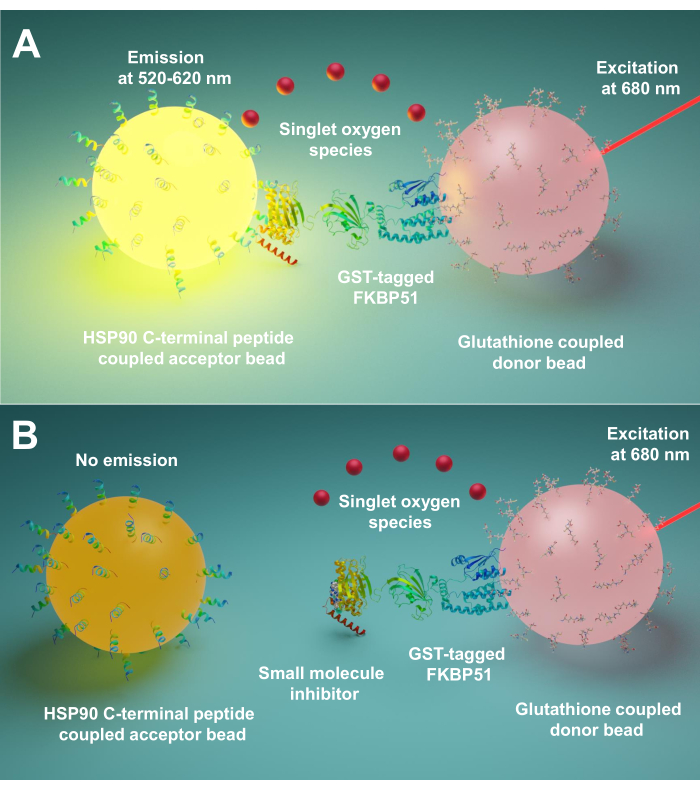
Figuur 1: Het basisprincipe van deze test. (A) Purified GST-FKBP51 interageert met glutathion-gebonden donorkralen. De 10 aminozuur lange peptiden die overeenkomen met de extreme C-terminus van Hsp90 zijn bevestigd aan acceptor kralen. De interactie tussen van Hsp90 afgeleide peptiden en het TPR-domein van FKBP51 brengt de donor- en acceptorparels in de nabijheid. Bij 680 nm licht een foto-ensitizer in de donorkraal op en zet zuurstof om in singletzuurstof. Het thioxeenderivaat op de acceptorkraal reageert met de singletzuurstof en genereert chemiluminescentie bij 370 nm. Deze energie activeert verder fluoroforen in dezelfde acceptorkraal om licht uit te stralen bij 520-620 nm. BWanneer kleine moleculen de interacties tussen Hsp90 en FKBP51 remmen, kunnen de donor- en acceptorparels de nabijheid niet bereiken. Dan vervalt de singletzuurstof met een korte levensduur en wordt er geen detecteerbaar signaal geproduceerd. Klik hier om een grotere versie van deze figuur te bekijken.
