Modellering av amyotrofisk lateralskleros (ALS) i laboratoriet görs unikt utmanande på grund av den överväldigande sporadiska karaktären hos över 80 % av fallen1, i kombination med det stora antalet genetiska mutationer som är kända för att vara sjukdomsorsakande2. Trots detta delar alla fall av ALS den förenande egenskapen att före direkt neuronal degeneration finns det dysfunktionell kommunikation mellan presynaptiska motorneuroner och postsynaptiska muskelceller3,4. Kliniskt, när patienter förlorar anslutningen av de återstående övre och nedre motorneuronerna, presenterar de funktioner av neuronal hyper- och hypoexcitabilitet under hela sjukdomen5,6,7,8,9, vilket återspeglar komplexa underliggande molekylära förändringar i dessa synapser, som vi som ALS-forskare försöker förstå.
Flera transgena modeller har illustrerat att försämring och desorganisation av den neuromuskulära korsningen sker med uttrycket av ALS-orsakande genetiska mutationer, inklusive SOD110, FUS11,12, C9orf7213,14,15,16 och TDP4317,18,19 genom morfologiska bedömningar, inklusive utvärdering av synaptiska boutons, ryggradstätheter och pre/postsynaptisk organisation. Mekaniskt, sedan Coles, Hodgkins och Huxleys landmärkespapper på 1930-talet, har det också varit möjligt att utvärdera synaptiska svar genom elektrofysiologiska tekniker i antingen in vitro-cellodling eller vävnadsskivberedningar20. Genom dessa strategier har många modeller av ALS visat synaptiska överföringsunderskott. Till exempel orsakar en mutant variant av TDP43 förbättrad avfyrningsfrekvens och minskar tröskeln för åtgärdspotential i NSC-34 (ryggmärg x neuroblastom hybridcellinje 34) motorneuronliknande celler21. Samma variant orsakar också dysfunktionell synaptisk överföring vid den neuromuskulära korsningen (NMJ) före uppkomsten av beteendemässiga motoriska underskott i en musmodell22. Det har tidigare visats att muterat FUS-uttryck resulterar i minskad synaptisk överföring vid NMJ i en drosophilamodell av FUS-ALS före rörelsedefekter11. En nyligen genomförd rapport med användning av inducerade pluripotenta stamceller härledda från C9orf72-expansionsbärare avslöjade en minskning av den lätt frisläppbara poolen av synaptiska vesiklar23. Sammantaget belyser dessa studier och andra vikten av att bygga en mer omfattande förståelse för mekanismerna bakom synaptisk signalering i sjukdomsrelevanta modeller av ALS. Detta kommer att vara avgörande för att förstå patobiologin för ALS och utveckla potentiella terapeutiska mål för patienter.
Metoder för ström- och spänningsklämningsceller har varit ovärderliga för att bestämma membranegenskaper såsom konduktans, vilande membranpotential och kvantinnehåll i enskilda synapser20,24. En av de betydande begränsningarna med elektrofysiologi är dock att den är tekniskt utmanande och bara ger insikter från en enda neuron åt gången. Konfokalmikroskopi med levande celler, i kombination med specifika fluorescerande sonder, ger möjlighet att undersöka den synaptiska överföringen av neuroner på ett spatiotemporalt sätt25,26,27. Även om det inte är ett direkt mått på neuronal excitabilitet, kan denna fluorescensmetod ge en relativ mätning av två molekylära korrelationer av synaptisk funktion: synaptisk vesiklarsättning och kalciumtransienter vid synaptiska terminaler.
När en åtgärdspotential når den presynaptiska terminala regionen av neuroner utlöses kalciumtransienter, vilket underlättar övergången från en elektrisk signal till processen för frisättning av neurotransmittorer28. Spänningsgrindade kalciumkanaler lokaliserade till dessa områden reglerar kalciumsignaleringen tätt för att modulera kinetiken för frisättning av neurotransmittorer29. De första rapporterade fluorescensbaserade inspelningarna av kalciumtransienter utfördes med användning av antingen dubbelvåglängdsindikatorn Fura-2 AM eller det enda våglängdsfärgämnet Fluo-3 AM30,31,32. Även om dessa färgämnen erbjöd stor ny insikt vid den tiden, lider de av flera begränsningar, såsom icke-specifik uppdelning i celler, aktiv eller passiv färgförlust från märkta celler, fotoblekning och toxicitet om de avbildas under längre tidsperioder33. Under det senaste decenniet har genetiskt kodade kalciumindikatorer blivit arbetshästar för avbildning av olika former av neuronal aktivitet. Dessa indikatorer kombinerar ett modifierat fluorescerande protein med ett kalciumkelatorprotein som snabbt växlar fluorescensintensitet efter bindningen av Ca2+ joner34. Tillämpningen av dessa nya indikatorer är omfattande, vilket möjliggör mycket enklare visualisering av intracellulära kalciumtransienter både in vitro– och in vivo-inställningar. En familj av dessa genetiskt kodade reportrar, känd som GCaMP, används nu i stor utsträckning. Dessa indikatorer innehåller en C-terminal kalmodulindomän, följt av grönt fluorescerande protein (GFP), och är begränsade av en N-terminal kalmodulinbindande region35,36. Kalciumbindning till kalmodulindomänen utlöser en interaktion med den kalmodulinbindande regionen, vilket resulterar i en konformationsförändring i den totala proteinstrukturen och en väsentlig ökning av fluorescensen hos GFP-delen35,36. Under årens lopp har denna familj av reportrar genomgått flera utvecklingar för att möjliggöra distinkta avläsningar för specifika kalciumtransienter med specifik kinetik (långsam, medium och snabb), var och en med något olika egenskaper37,38. Här har användningen av reportern GcaMP6 belysts, som tidigare har visat sig detektera enstaka åtgärdspotentialer och dendritiska kalciumtransienter i neuroner både in vivo och in vitro37.
Kalciumtransienter i den presynaptiska regionen utlöser synaptiska vesiklarfusionshändelser, vilket orsakar frisättning av neurotransmittorer i synapsen och initiering av signalhändelser i den postsynaptiska cellen28,39. Synaptiska vesiklar frigörs både snabbt och återvinns, eftersom cellen homeostatiskt upprätthåller en stabil cellmembranyta och lätt frigörbar pool av fusionskapabla membranbundna vesiklar40. Styrylfärgämnet som används här har en affinitet mot lipidmembran och ändrar specifikt dess emissionsegenskaper baserat på ordningen av den omgivande lipidmiljön41,42. Således är det ett idealiskt verktyg för märkning av återvinning av synaptiska vesiklar och efterföljande spårning av dessa vesiklar när de senare frigörs efter neuronal stimulering41,42. Protokollet som har genererats och optimerats är en anpassning av de begrepp som ursprungligen beskrevs av Gaffield och kollegor, vilket gör att vi kontinuerligt kan visualisera styrylfärgmärkt synaptisk vesiklarecta över tiden41.
Här beskrivs två relaterade fluorescensbaserade metoder som på ett tillförlitligt sätt rapporterar specifika cellulära händelser som är involverade i synaptisk överföring. Protokoll har definierats för att undersöka dynamiken i depolariseringsmedierad presynaptisk terminal kalciuminflöde och synaptisk vesikleexocytos i odlade neuroner. Här fokuseras metoder och representativa resultat på att använda primära gnagare kortikala eller motoriska neuroner som in vitro-modellsystem, eftersom det finns publicerade studier med dessa celltyper43,44. Dessa metoder är emellertid också tillämpliga på differentierade humana i3 kortikala nervceller45, eftersom vi också har haft framgång med båda protokollen i för närvarande pågående experiment i vårt laboratorium. Det allmänna protokollet beskrivs i ett stegvis linjärt format, som visas i figur 1. I korthet, för att studera kalciumdynamiken i neuriter transfekteras mogna neuroner med plasmid-DNA för att uttrycka den fluorescerande reportern GCaMP6m under en Cytomegalovirus (CMV) promotor37,46. Transfekterade celler har en låg nivå av basal grön fluorescens, vilket ökar i närvaro av kalcium. Regioner av intresse specificeras för att övervaka fluorescensförändringar under hela vår manipulation. Detta gör det möjligt att mäta mycket rumsligt och tidsmässigt lokaliserade fluktuationer i kalcium37,46. För utvärdering av synaptisk vesiklarfusion och frisättning laddas mogna neuroner med styrylfärgämne införlivat i synaptiska vesiklarmembran när de återvinns, reformeras och laddas om med neurotransmittorer i presynaptiska celler41,42,43,47,48. De nuvarande färgämnena som används för detta ändamål märker synaptiska vesiklar längs neuriter och används som proxy för dessa regioner i live-imaging-experiment, vilket visades genom samfärgning av styrylfärgämne och synaptotagmin av Kraszewski och kollegor49. Här ingår representativa bilder av liknande färgning som också har utförts (figur 2A). Tidigare utredare har i stor utsträckning använt sådana färgämnen för att rapportera synaptisk vesikeldynamik vid den neuromuskulära korsningen och hippocampala neuroner48,49,50,51,52,53,54,55,56 . Genom att välja punkterade regioner av färgladdade vesiklar och genom att övervaka minskningar av fluorescensintensiteten efter vesiklarsättning kan funktionell synaptisk överföringskapacitet och temporal frisättningsdynamik studeras efter stimulering43. För båda metoderna används ett medium innehållande en hög koncentration av kaliumklorid för att depolarisera celler för att efterlikna neuronal aktivitet. Bildparametrar anges för att fånga intervall under sekunden som spänner över en baslinjenormalisering följt av vår stimuleringsfångstperiod. Fluorescensmätningar vid varje tidpunkt bestäms, normaliseras till bakgrunden och kvantifieras under den experimentella tidsperioden. Kalciuminflödesmedierad GCaMP6m fluorescensökning eller effektiv synaptisk vesiklarexocytocytos styrylfärgämne frisättning fluorescens minskning kan detekteras genom denna strategi. Detaljerad metodologisk inställning och parametrar för dessa två protokoll och en diskussion om deras fördelar och begränsningar beskrivs nedan.
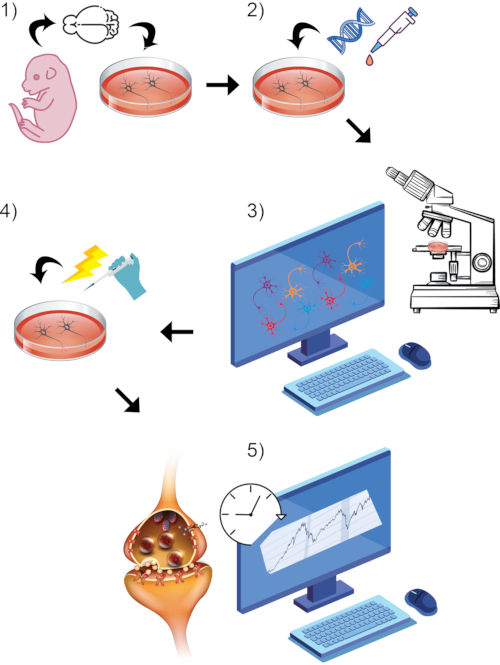
Bild 1: Visuell återgivning av den övergripande allmänna protokollprocessen. (1) Isolera och odla primära gnagareneuroner in vitro till vald mognadstidspunkt. (2) Introducera GCaMP-DNA eller styrylfärgämne som rapportörer av synaptisk aktivitet. (3) Inställningsavbildningsparadigm med hjälp av live-imaging-utrustat konfokalmikroskop och tillhörande programvara. Påbörja baslinjeinspelningsperioden. (4) Medan celler fortfarande genomgår live-image-fångst, stimulera neuroner via hög KCl-badperfusion. (5) Bedöma fluorescensintensitetsmätningar över tid för att mäta kalciumtransienter eller synaptisk vesiklarfusion. Klicka här för att se en större version av denna figur.
