La modélisation de la sclérose latérale amyotrophique (SLA) en laboratoire est rendue particulièrement difficile en raison de la nature extrêmement sporadique de plus de 80 % des cas1, associée au grand nombre de mutations génétiques connues pour être à l’origine de la maladie2. Malgré cela, tous les cas de SLA partagent la caractéristique unificatrice qu’avant la dégénérescence neuronale pure et simple, il existe une communication dysfonctionnelle entre les motoneurones présynaptiques et les cellules musculaires postsynaptiques3,4. Cliniquement, à mesure que les patients perdent la connectivité des motoneurones supérieurs et inférieurs restants, ils présentent des caractéristiques d’hyper- et d’hypoexcitabilité neuronale tout au long de la maladie5,6,7,8,9, reflétant des changements moléculaires sous-jacents complexes à ces synapses, que nous, en tant que chercheurs sur la SLA, cherchons à comprendre.
Plusieurs modèles transgéniques ont montré que la détérioration et la désorganisation de la jonction neuromusculaire se produisent avec l’expression de mutations génétiques responsables de la SLA, y compris SOD110, FUS11,12, C9orf7213,14,15,16 et TDP4317,18,19 par des évaluations morphologiques, y compris l’évaluation des boutons synaptiques, des densités de la colonne vertébrale et de l’organisation pré/postsynaptique. Mécaniquement, depuis les articles historiques de Cole, Hodgkin et Huxley dans les années 1930, il a également été possible d’évaluer les réponses synaptiques grâce à des techniques électrophysiologiques dans des cultures cellulaires in vitro ou des préparations de tranches de tissu20. Grâce à ces stratégies, de nombreux modèles de SLA ont démontré des déficits de transmission synaptique. Par exemple, une variante mutante de TDP43 améliore la fréquence de tir et diminue le seuil de potentiel d’action dans les cellules de type moteur-neurone NSC-34 (moelle épinière x neuroblastome hybride lignée 34)21. Cette même variante provoque également une transmission synaptique dysfonctionnelle à la jonction neuromusculaire (NMJ) avant l’apparition de déficits moteurs comportementaux chez un modèle murin22. Il a déjà été démontré que l’expression mutante de FUS entraîne une réduction de la transmission synaptique au NMJ dans un modèle de drosophile de FUS-ALS avant les défauts locomoteurs11. Un rapport récent utilisant des cellules souches pluripotentes induites dérivées de porteurs d’expansion C9orf72 a révélé une réduction du pool facilement libérable de vésicules synaptiques23. Dans l’ensemble, ces études et d’autres soulignent l’importance d’acquérir une compréhension plus complète des mécanismes sous-jacents à la signalisation synaptique dans les modèles de SLA pertinents pour la maladie. Cela sera essentiel pour comprendre la pathobiologie de la SLA et développer des cibles thérapeutiques potentielles pour les patients.
Les méthodes de serrage du courant et de la tension ont été inestimables pour déterminer les propriétés de la membrane telles que la conductance, le potentiel de la membrane au repos et le contenu quantifial des synapses individuelles20,24. Cependant, l’une des limites importantes de l’électrophysiologie est qu’elle est techniquement difficile et ne fournit des informations qu’à partir d’un seul neurone à la fois. La microscopie confocale à cellules vivantes, associée à des sondes fluorescentes spécifiques, offre la possibilité d’étudier la transmission synaptique des neurones de manière spatio-temporelle25,26,27. Bien qu’il ne s’agisse pas d’une mesure directe de l’excitabilité neuronale, cette approche de fluorescence peut fournir une mesure relative de deux corrélations moléculaires de la fonction synaptique: la libération de vésicules synaptiques et les transitoires calciques aux terminaux synaptiques.
Lorsqu’un potentiel d’action atteint la région terminale présynaptique des neurones, des transitoires calciques sont déclenchés, facilitant la transition d’un signal électrique au processus de libération de neurotransmetteurs28. Les canaux calciques voltage-dépendants localisés dans ces zones régulent étroitement la signalisation du calcium pour moduler la cinétique de libération de neurotransmetteurs29. Les premiers enregistrements basés sur la fluorescence des transitoires de calcium ont été effectués à l’aide de l’indicateur à double longueur d’onde Fura-2 AM ou du colorant à longueur d’onde unique Fluo-3 AM30,31,32. Bien que ces colorants aient offert de nouvelles perspectives à l’époque, ils souffrent de plusieurs limitations telles que le compartimentage non spécifique au sein des cellules, la perte de colorant actif ou passif des cellules marquées, le photoblanchiment et la toxicité s’ils sont imagés sur de longues périodes de temps33. Au cours de la dernière décennie, les indicateurs de calcium génétiquement codés sont devenus les chevaux de bataille pour l’imagerie de diverses formes d’activité neuronale. Ces indicateurs combinent une protéine fluorescente modifiée avec une protéine chélateur de calcium qui change rapidement d’intensité de fluorescence après la liaison des ions Ca2+34. L’application de ces nouveaux indicateurs est vaste, permettant une visualisation beaucoup plus facile des transitoires de calcium intracellulaires à la fois in vitro et in vivo. Une famille de ces rapporteurs génétiquement codés, connue sous le nom de GCaMP, est maintenant largement utilisée. Ces indicateurs contiennent un domaine de calmoduline C-terminal, suivi d’une protéine fluorescente verte (GFP), et sont coiffés d’une région de liaison à la calmoduline N-terminale35,36. La liaison du calcium au domaine de la calmoduline déclenche une interaction avec la région de liaison à la calmoduline, entraînant un changement conformationnel de la structure protéique globale et une augmentation substantielle de la fluorescence de la fraction GFP35,36. Au fil des ans, cette famille de rapporteurs a subi plusieurs évolutions pour permettre des lectures distinctes pour des transitoires de calcium particuliers avec une cinétique spécifique (lente, moyenne et rapide), chacune avec des propriétés légèrement différentes37,38. Ici, l’utilisation du rapporteur GcaMP6 a été mise en évidence, qui a déjà été montré pour détecter les potentiels d’action unique et les transitoires de calcium dendritique dans les neurones à la fois in vivo et di vitro37.
Les transitoires calciques dans la région présynaptique déclenchent des événements de fusion des vésicules synaptiques, provoquant la libération de neurotransmetteurs dans la synapse et l’initiation d’événements de signalisation dans la cellule postsynaptique28,39. Les vésicules synaptiques sont à la fois rapidement libérées et recyclées, car la cellule maintient homéostatiquement une surface de membrane cellulaire stable et un pool facilement libérable de vésicules membranaires capables de fusion40. Le colorant styryl utilisé ici a une affinité pour les membranes lipidiques et modifie spécifiquement ses propriétés d’émission en fonction de l’ordre de l’environnement lipidique environnant41,42. C’est donc un outil idéal pour marquer les vésicules synaptiques de recyclage et le suivi ultérieur de ces vésicules telles qu’elles sont libérées plus tard après une stimulation neuronale41,42. Le protocole qui a été généré et optimisé est une adaptation des concepts décrits initialement par Gaffield et ses collègues, ce qui nous permet de visualiser en continu les puncta de vésicules synaptiques marquées par un colorant styryl au fil du temps41.
Ici, deux méthodologies connexes basées sur la fluorescence sont décrites, rapportant de manière fiable des événements cellulaires spécifiques impliqués dans la transmission synaptique. Des protocoles ont été définis pour sonder la dynamique de l’afflux de calcium terminal présynaptique médié par la dépolarisation et de l’exocytose des vésicules synaptiques dans les neurones en culture. Ici, les méthodes et les résultats représentatifs sont axés sur l’utilisation de corticaux ou de motoneurones primaires de rongeurs comme système de modèle in vitro, car il existe des études publiées utilisant ces types de cellules43,44. Cependant, ces méthodes sont également applicables aux neurones corticaux i3 humains différenciés45, car nous avons également eu du succès avec les deux protocoles dans l’expérimentation actuellement en cours dans notre laboratoire. Le protocole général est décrit dans un format linéaire par étapes, illustré à la figure 1. En bref, pour étudier la dynamique du calcium dans les neurites, les neurones matures sont transfectés avec de l’ADN plasmidique pour exprimer le rapporteur fluorescent GCaMP6m sous un promoteur de cytomégalovirus (CMV)37,46. Les cellules transfectées ont un faible niveau de fluorescence vert basal, ce qui augmente en présence de calcium. Les régions d’intérêt sont spécifiées pour surveiller les changements de fluorescence tout au long de notre manipulation. Cela permet de mesurer des fluctuations de calcium très localisées dans l’espace et dans le temps37,46. Pour évaluer la fusion et la libération des vésicules synaptiques, les neurones matures sont chargés de colorant styryl incorporé dans les membranes des vésicules synaptiques au fur et à mesure qu’ils sont recyclés, reformés et rechargés en neurotransmetteurs dans les cellules présynaptiques41,42,43,47,48. Les colorants actuels utilisés à cette fin marquent les vésicules synaptiques le long des neurites et sont utilisés comme proxy pour ces régions dans des expériences d’imagerie en direct, comme l’ont montré la co-coloration du colorant styryl et de la synaptotagmine par Kraszewski et ses collègues49. Sont incluses ici des images représentatives de taches similaires qui ont également été effectuées (Figure 2A). Des chercheurs antérieurs ont largement utilisé ces colorants pour rapporter la dynamique des vésicules synaptiques à la jonction neuromusculaire et aux neurones de l’hippocampe48,49,50,51,52,53,54,55,56 . En sélectionnant les régions ponctuées des vésicules chargées de colorant et en surveillant les diminutions de l’intensité de fluorescence après la libération des vésicules, la capacité de transmission synaptique fonctionnelle et la dynamique temporelle de libération peuvent être étudiées après stimulation43. Pour les deux méthodes, un milieu contenant une forte concentration de chlorure de potassium est utilisé pour dépolariser les cellules afin d’imiter l’activité neuronale. Les paramètres d’imagerie sont spécifiés pour capturer des intervalles inférieurs à la seconde couvrant une normalisation de base suivie de notre période de capture de stimulation. Les mesures de fluorescence à chaque point temporel sont déterminées, normalisées à l’arrière-plan et quantifiées au cours de la période expérimentale. L’augmentation de la fluorescence GCaMP6m médiée par l’afflux de calcium ou la diminution efficace de l’exocytose de la vésicule synaptique par la libération de colorant styryl peut être détectée grâce à cette stratégie. La configuration méthodologique détaillée et les paramètres de ces deux protocoles ainsi qu’une discussion sur leurs avantages et leurs limites sont décrits ci-dessous.
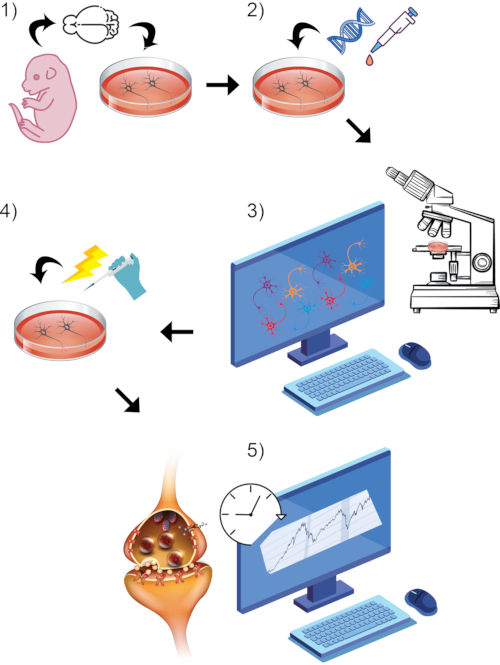
Figure 1 : Rendu visuel de l’ensemble du processus général du protocole. (1) Isoler et cultiver les neurones primaires des rongeurs in vitro jusqu’au point de temps de maturation choisi. (2) Introduire l’ADN GCaMP ou le colorant styryl comme rapporteurs de l’activité synaptique. (3) Configurer le paradigme de l’imagerie à l’aide d’un microscope confocal équipé d’imagerie en direct et du logiciel associé. Commencez la période d’enregistrement de référence. (4) Pendant que les cellules subissent encore une capture d’images en direct, stimulez les neurones via une perfusion de bain KCl élevée. (5) Évaluer les mesures d’intensité de fluorescence au fil du temps pour mesurer les transitoires calciques ou la fusion des vésicules synaptiques. Veuillez cliquer ici pour voir une version agrandie de cette figure.
