Selective Ribosome Profiling (SeRP) è l’unico metodo, ad oggi, che cattura e caratterizza le interazioni co-traslazionali, in vivo, in modo diretto 1,2,3,4,5,6. SeRP consente la profilazione globale delle interazioni di qualsiasi fattore con la traduzione dei ribosomi nella risoluzione vicino al codone 2,7.
Il metodo si basa sul congelamento flash delle cellule in crescita e sulla conservazione della traduzione attiva. I lisati cellulari vengono quindi trattati con RNasi I per digerire tutto l’mRNA nella cellula ad eccezione dei frammenti di mRNA protetti dai ribosomi chiamati “impronte di ribosomi”. Il campione viene quindi diviso in due parti; una parte viene utilizzata direttamente per l’isolamento di tutte le impronte ribosomiali cellulari, rappresentando tutta la traduzione in corso nella cellula. La seconda parte viene utilizzata per l’affinità-purificazione del sottoinsieme specifico di ribosomi associati a un fattore di interesse, ad esempio: modifica di enzimi, fattori di traslocazione, chaperoni di piegatura e interazioni complesso-assemblaggio. Le impronte ribosomiali purificate dall’affinità sono collettivamente chiamate interattoma. Quindi, gli mRNA protetti dai ribosomi vengono estratti e utilizzati per la generazione di librerie di cDNA, seguiti dal sequenziamento profondo.
L’analisi comparativa dei campioni totali di translatome e interactome consente l’identificazione di tutti gli orfs che si associano al fattore di interesse, nonché la caratterizzazione di ciascun profilo di interazione orf. Questo profilo riporta le precise sequenze di insorgenza e terminazione dell’incarico da cui si possono dedurre i codoni decodificati e i rispettivi residui della catena polipeptidica emergente, nonché sulle variazioni di velocità del ribosoma durante l’interazione 7,8. La Figura 1 illustra il protocollo come schematico.
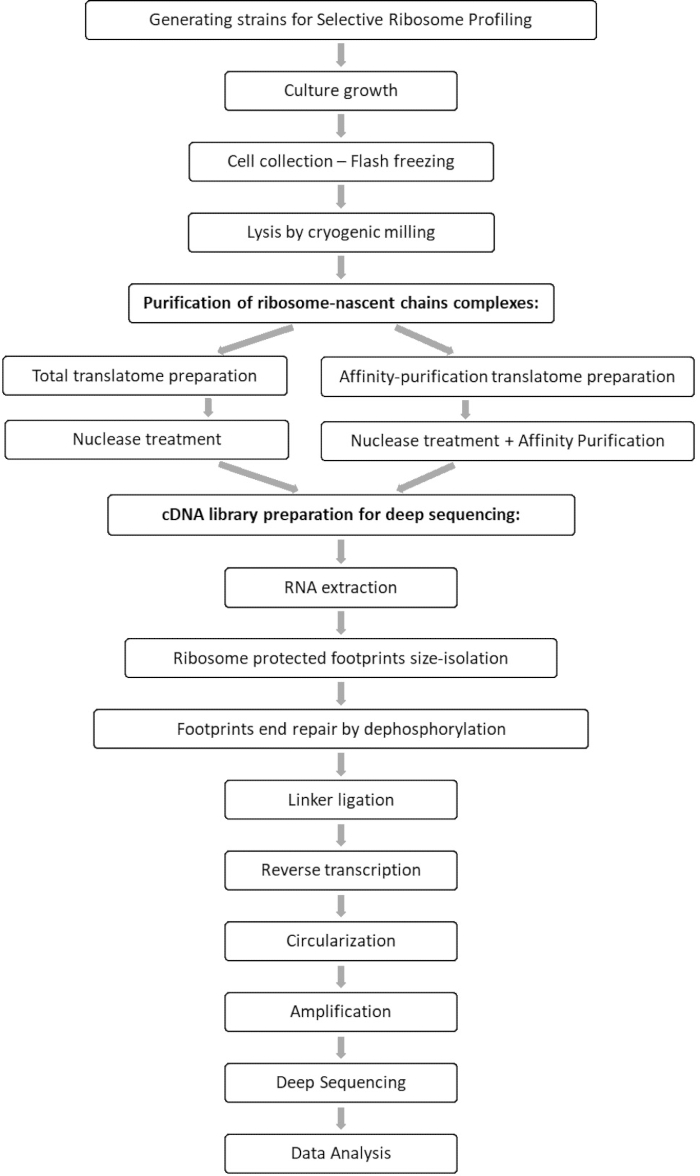
Figura 1: Panoramica del protocollo SeRP. Questo protocollo può essere eseguito nella sua interezza entro 7-10 giorni. Fare clic qui per visualizzare una versione più grande di questa figura.
