All experiments were performed in accordance with The Ethics Committee of the Faculty of Philosophy, Sciences, and Letters of Ribeirão Preto (CEUA/FFCLRP 18.5.35.59.5).
1. 6-OHDA lesion
- Use Sprague-Dawley male rats weighing 200-250 g at the beginning of the experiments (6 weeks). House the animals (2-3 per cage) under standard laboratory conditions (12:12 h light/dark cycle, lights on at 06:00 h, temperature-controlled facilities (22-24 °C), with food and water available ad libitum.
NOTE: A complete description of how to generate severe 6-OHDA-lesioned rats is described elsewhere42. - Appropriately acclimate animals before the beginning of any experimental procedure.
- Administer (intraperitoneally, i.p.) the norepinephrine transporter inhibitor imipramine (20 mg/kg, dissolved in 0.9% saline solution) 30 min before surgery.
NOTE: Imipramine administration will increase 6-OHDA selectivity for dopaminergic neurons. - Administer ketamine/xylazine (70/10 mg/kg) intraperitoneally.
- Make sure the animal is deeply anesthetized by the lack of response to toe pinch and positioned in a prone position in the stereotaxic apparatus on the top of a heating pad.
- Remove the fur on the head and disinfect the surgical area 3 times, alternating between an iodine-based scrub and alcohol.
- Use a scalpel to make an incision (approximately 1 cm) at the region where the microinjection will occur.
- Clean the skull region with cotton swabs and ensure the Bregma and Lambda are exposed.
- Ensure MFB stereotaxic coordinates are taken from bregma43,44,45: -4.3 mm anterior, 1.6 mm lateral (right side), and 8.3 mm ventral (from the dura mater).
- Administer the 6-OHDA at a rate of 0.4 µL/min (10 µg in 4 µL of saline solution containing 0.1% ascorbic acid) unilaterally in the right medial forebrain bundle (MFB) using a 50 µL Hamilton glass syringe.
- At the end of the surgery, suture the scalp incision and rehydrate the animal with warm (~37 °C) sterile 0.9% saline solution (~10 mL/kg, s.c.).
- Remove the animal from the stereotaxic frame and place it in a warmed recovery cage. Monitor until consciousness is regained.
- Assess the effectiveness of the dopaminergic lesion 4 weeks post-lesion using a stepping test46,47.
NOTE: In this test, the akinesia of the forelimb contralateral to the lesion is assessed through the number of adjusting steps with the forepaw contralateral to the lesion. Rats presenting three or fewer adjusting steps with the contralateral forelimb are included in the study as putative severely 6-OHDA-lesioned rats48.
2. L-DOPA chronic treatment
- Start the chronic treatment on Monday, 4 weeks post-6-OHDA lesion.
NOTE: Since AIMs will be videotaped for 3 consecutive days per week (Wednesday, Thursday, and Friday) for 3 weeks, it is recommended to start the treatment on Monday. - Treat the rats with freshly prepared L-DOPA (via subcutaneous (s.c.), 1 mL/kg) plus benserazide hydrochloride (5 mg/kg and 12.5 mg/kg, respectively) for 3 weeks, once daily from Monday to Friday45.
NOTE: Once established, AIMs will manifest with each administration of L-DOPA. Therefore, L-DOPA administration 2-4 times a week after the 3-week induction period is sufficient to maintain a stable expression of AIMs48.
3. AIMs recording and scoring
- Perform experiments between 09:00 h and 17:00 h. Ensure that the investigator is entirely unaware of the rats' identity and pharmacological treatment.
- Carefully put the rat inside a transparent cylinder (20 cm diameter x 40 cm tall) and allow it to acclimatize for at least 15 min. Ensure that the floor is covered with the bedding material. Mirrors must be positioned behind the cylinder so the experimenter can observe the animal from all possible angles.
NOTE: If there is more than one rat per experiment, use a cylinder for each rat. Manipulation of the animals between sessions can interfere with the behavioral analysis. - Position a high-resolution video camera in a way that favors the viewing of axial, limb, and orolingual AIMs. Since rotational behavior is present under these experimental conditions, the mirrors behind the cylinder will allow tracking AIMs at a 360° angle. Use a tripod or fix the camera directly to the bench.
NOTE: Orolingual AIMs can be challenging to score, especially if there is a high incidence of rotational behavior. Capturing videos with the camera positioned slightly below the plane where the animals are at a 15° angle will help observe this movement subtype. - Gently remove the animal from the cylinder and administer L-DOPA (5 mg/kg combined with 12.5 mg/kg of benserazide, s.c.).
- Put the animal back in the cylinder and start a timer to track AIMs after L-DOPA injection. Use a video camera to record AIMs for 180 min after the injection of L-DOPA. Although the analysis can be carried out by direct observation of the animal, offline scoring is preferred.
NOTE: Ensure the timer is also videotaped so that AIMs can be scored at precise times after L-DOPA administration. - Score AIMs at 30 min intervals up to 180 min post-L-DOPA injection (see Table 1 for definitions) as initially described20,23. Scores must be given over 1-2 min epochs and classified as axial, limb, or orolingual (Figure 1).
- Carefully observe each AIM and attribute scores for severity and amplitude. Use Table 1 for instructions. Do not include normal behaviors, such as rearing, sniffing, grooming, and gnawing in the rating. There will be a total of six observation periods (30, 60, 90, 120, 150, and 180 min).
| SUBTYPES |
| Axial: torsion of head, neck, and trunk |
| Limb: involuntary movements of both distal and proximal forelimb |
| Orolingual: involuntary movements of the orofacial muscles |
| SEVERITY |
| 0: Absent |
| 1: Occasional (with interruptions, present in less than half of the observation time) |
| 2: Frequent (with interruptions but present in more than half of the observation time) |
| 3: Continuous but interrupted by external sensory stimuli |
| 4: Continuous and not interrupted by external sensory stimuli |
| AMPLITUDE |
| Axial |
| 1: Torsion of head and neck at approximately 30° angle |
| 2: Torsion of head and neck at approximately 30° < angle ≤ 60° |
| 3: Torsion of the head, neck, and upper trunk at approximately 60° < angle ≤ 90° |
| 4: Torsion of head, neck, and trunk at > 90° angle, often causing the rat to lose balance |
| Limb |
| 1: Small involuntary movements of the distal forelimb |
| 2: Movements of low amplitude causing translocation of both distal and proximal forelimb |
| 3: Involuntary movements of the whole limb, including shoulder muscles |
| 4: Strong limb and shoulder movements, often similar to ballism |
| Orolingual |
| 1: Small involuntary movements of the orofacial muscles |
| 2: Orofacial movements of high amplitude with tongue protrusion |
Table 1: AIMS definition and rating criteria.

Figure 1: Sequence of photos showing each AIM subtype. (A to A'") Axial AIM showing dystonic-like movements in different angles (A: 30° angle, A': 30° < angle ≤ 60°, A": 60° < angle ≤ 90°, and A'": > 90° angle). (B to B") Limb AIM (black arrows) showing involuntary movements of the distal (B and B') and whole forelimb (including shoulder, B"). (C) Orolingual AIM (red arrow) of high amplitude with tongue protrusion. Please click here to view a larger version of this figure.
4. Data analysis
- Compute axial, limb, or orolingual AIMs subtypes by multiplying the severity and amplitude scores for each observation period after L-DOPA administration (i.e., six observation periods) as previously described20. The theoretical maximum score that one animal could accumulate in one testing session is 240 (maximum score per observation period is 40, see Table 1 for details).
- Plot AIMs scores for all the six observation periods recorded on Wednesday, Thursday, and Friday over the 3 weeks of chronic L-DOPA administration (Figure 2). AIMs can be summed up to indicate each animal's maximum score on each scoring day (Figure 3) or each week (Figure 4) over the 3 weeks of chronic L-DOPA administration.
- Verify if the data has a close-to-normal distribution before selecting the statistical analysis approach.
NOTE: AIMs rating scales consist of ordinal data and can be analyzed with nonparametric statistics. Parametric tests are widely used when comparing AIMs scores across experimental groups following repeated testing sessions40.
Although the AIMs patterns observed in rats are simpler and limited compared to those observed in humans and nonhuman primates, this model reproduces both hyperkinetic and dystonic-like movements induced by chronic L-DOPA administration. Here we present data collected from a group (n = 10) of unilaterally 6-OHDA-lesioned rats chronically treated with L-DOPA (5 mg/kg combined with 12.5 mg/kg of benserazide) for 3 weeks (Monday to Friday). Note that the data presented in Figure 2, Figure 3, and Figure 4 are from the same animals.
Figure 2 shows the time course analysis for scores applied to axial, limb, and orolingual AIMs over 3 weeks of chronic L-DOPA administration. Each AIM score subtype at a specific observation period is reported as the multiplication of severity and amplitude scores in this analysis. This type of analysis can be convenient to explore the effect of drugs that interfere with the temporal profile of AIMs.Note that L-DOPA-induced peak-dose dyskinesias occur between 30-90 min with a gradual decrease after 120 min post-injection (Figure 2).
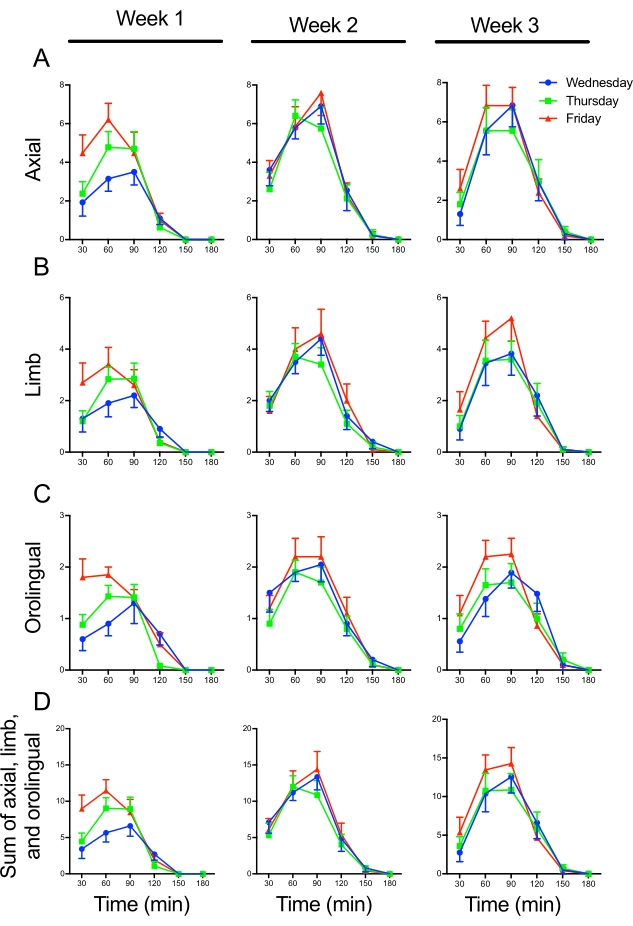
Figure 2: Time course analysis showing axial, limb, and orolingual AIMs over 3 weeks of chronic L-DOPA administration (5 mg/kg combined with 12.5 mg/kg of benserazide) to 6-OHDA-lesioned rats. (A) Axial, (B) limb, and (C) orolingual AIMs were scored on Wednesday, Thursday, and Friday over 3 weeks of chronic L-DOPA administration. Each AIM subtype is reported as the multiplication of severity times amplitude on each of the six observation periods. Each AIM subtype can be analyzed individually or summed up as indicated in (D). Note that the peak severity of the AIMs will gradually increase during the first week of chronic L-DOPA administration. Data are expressed as mean ± SEM (n = 10). Please click here to view a larger version of this figure.
Figure 3 shows the sum of AIMs scores presented for all the six observation periods depicted in Figure 2 (datasets are color-coded for easy comparisons). This analysis indicates each animal's maximum score on each scoring day (Wednesday, Thursday, and Friday) over the 3 weeks of chronic L-DOPA administration. This type of analysis can be advantageous to explore the effect of compounds with potential antidyskinetic profiles, especially because AIMs scores are stable over weeks 2 and 3 of chronic L-DOPA administration. Furthermore, weekly AIMs scores (Figure 4) can be generated from the sum of scores attributed to 6-OHDA-lesioned rats on Wednesday, Thursday, and Friday.
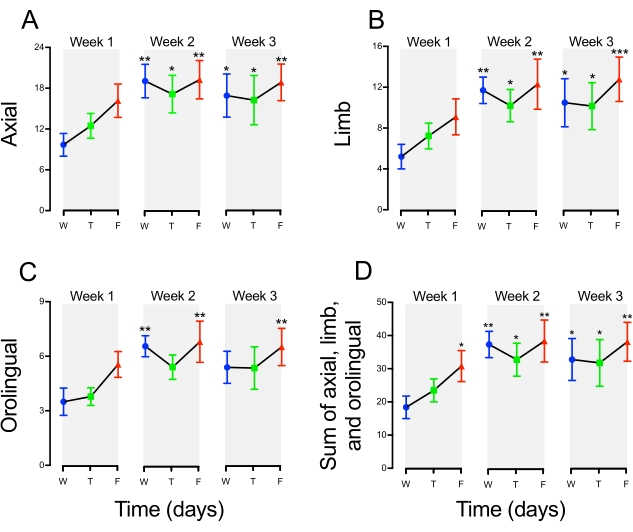
Figure 3: Sum of axial, limb, and orolingual AIMs on individual scoring days over 3 weeks of chronic L-DOPA administration (5 mg/kg combined with 12.5 mg/kg of benserazide) to 6-OHDA-lesioned rats. In this figure, AIMs scores presented in each of the six observation periods (see details in Figure 2 legend) were summed. Individual datasets were color-coded to compare with the data displayed in Figure 2. This analysis indicates each animal's maximum score on each scoring day (Wednesday – W, Thursday – T, and Friday – F) following 3 weeks of chronic L-DOPA administration. (A) Axial, (B) Limb, and (C) orolingual AIMs. (D) Sum of axial, limb, and orolingual AIMs. One-way repeated measures ANOVA and Holm-Sidak post-hoc test revealed that AIMs increase gradually over the chronic administration of L-DOPA. Note that AIMs scores are stable over weeks 2 and 3 of chronic L-DOPA administration (*p < 0.05, **p < 0.01, ***p < 0.001 vs. Wednesday (W) on Week 1). Data are expressed as mean ± SEM (n = 10). Please click here to view a larger version of this figure.
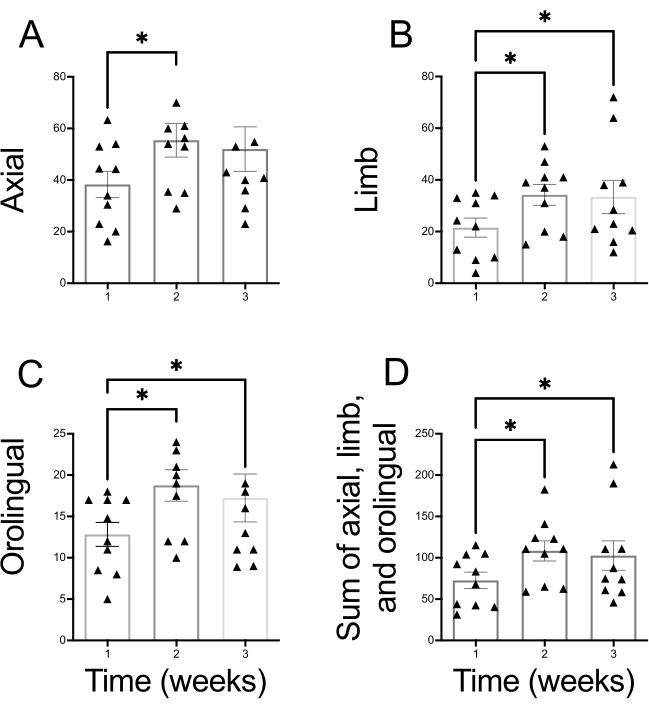
Figure 4: Sum of weekly axial, limb, and orolingual AIMs throughout chronic L-DOPA administration (5 mg/kg combined with 12.5 mg/kg of benserazide) to 6-OHDA-lesioned rats. In this figure, AIMs scored each week (Wednesday, Thursday, and Friday – see Figure 3) were summed. This analysis indicates each animal's maximum score on each week of chronic L-DOPA administration. (A) Axial, (B) Limb, and (C) orolingual AIMs. (D) Sum of axial, limb, and orolingual AIMs. One-way repeated measures ANOVA and Holm-Sidak post-hoc test revealed that AIMs increase over the chronic administration of L-DOPA (*p < 0.05 vs. Week 1). Data are expressed as mean ± SEM (n = 10). Please click here to view a larger version of this figure.
