The workflow described in Figure 1 was used to perform morpholino-based genetic perturbation at the zebrafish one-cell stage. Pigmentation analysis was performed using various methods, as mentioned below. To illustrate the representative results, standardized volumes of antisense morpholino targeting h2afv and ca14 genes were injected in the yolk or one-cell stage of the zebrafish embryo. The initial phenotyping based on brightfield imaging was performed at 48 hours post fertilization (hpf), when all the five pigmented melanophore stripes (dorsal, ventral, yolk, two lateral) were evident.
Figure 2A,B represent an approach to analyze pigmentation by calculating the number of lateral melanophores per embryo. Lateral melanophores were manually counted at the 48 hpf stage by zooming in on the lateral region of the tilted zebrafish. An alternative method to quantify the pigmented melanophores (Supplemental Figure S1) involves the manual counting of the head melanophores by imaging the dorsal view of the zebrafish.
The melanin content per embryo is calculated by measuring the mean gray value, keeping the region of interest constant with the help of ImageJ software. Figure 2C-F show that the mean gray value of ca14 and h2afv morphants was higher than in control morphants. As the mean gray value is inversely proportional to melanin content, knockdown of ca14 and h2afv genes led to a decrease in melanin content per embryo. Another robust method to quantify the melanin content is a NaOH-based spectrophotometric absorption method. As shown in Figure 2G, the total melanin content in ca14 morphants was less than in control morphants.
Fluorescent imaging revealed two different stages of zebrafish melanophore development: early specified melanophores (mitfa:gfp) and differentiating melanophores (ftyrp:gfp). The morpholino-based alteration was evaluated by fluorescent imaging (Figure 3A and Figure 3C). To further validate these observations, the frequency of GFP-positive cells was calculated using FACS. The knockdown of h2afv but not ca14 significantly reduced the number of melanophores with respect to control(Figure 3A-D). Moreover, a reduction in the mean fluorescence intensity/area can be calculated using ImageJ software, keeping the region of interest constant. H2afv morphants show a significant decrease in the mean fluorescent intensity/area relative to control morphants (Figure 3E,F).
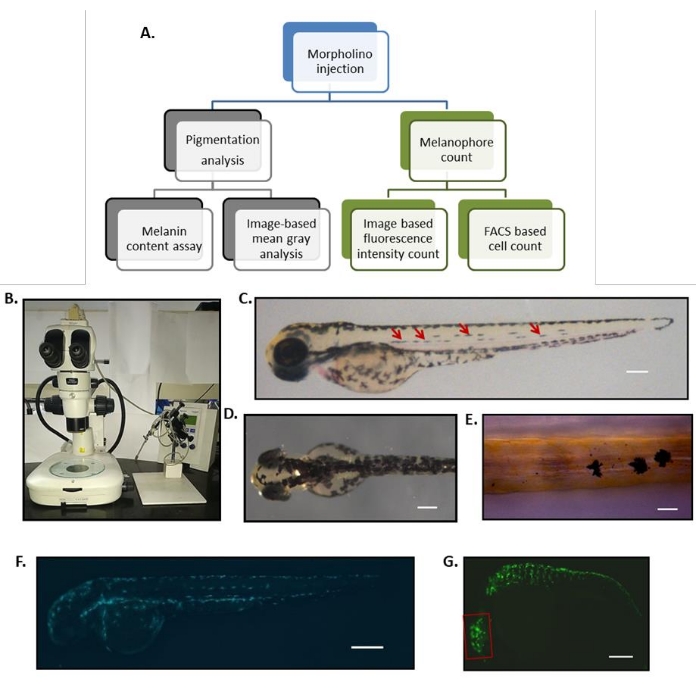
Figure 1: Workflow for a reverse genetic approach to identify regulators of melanocyte biology using zebrafish. (A) Flowchart depicting the experimental workflow starting from microinjection to evaluating pigmentation by various methods. (B) Microinjection needle holder and micromanipulator, along with a dissecting microscope, is a typical setup for microinjection of one-cell stage zebrafish embryos. (C) Embryonic melanocyte patterning in zebrafish at 3 dpf showing all five stripes: dorsolateral, two lateral midlines (pointed by four red arrows), ventral stripe, and yolk stripe. (D) To study the pigmentation outcome upon morpholino injection, lateral midline melanophores can be counted under a stereomicroscope. Brightfield image of a dorsal region at 2 dpf; melanin content can be quantified by measuring mean grey value. (E) The mean gray values are inversely proportional to the melanin content of the embryo. Melanin-filled melanophores at 2 dpf in wild-type embryos. (F) Melanin content assay can be performed to estimate the melanin production within these melanophores. Transgenic zebrafish labeling the cells expressing TYRP1 protein at 2 dpf. (G) Fluorescently labeled cells can be quantified using FACS; scale bar = 100 µm. Transgenic zebrafish labeling the cells expressing Mitfa protein at 1 dpf. Meanfluorescence intensity per animal can be measured using ImageJ software. Scale bars = 100 µm. Abbreviations: dpf = days post fertilization; FACS = fluorescence-activated cell sorting. Please click here to view a larger version of this figure.
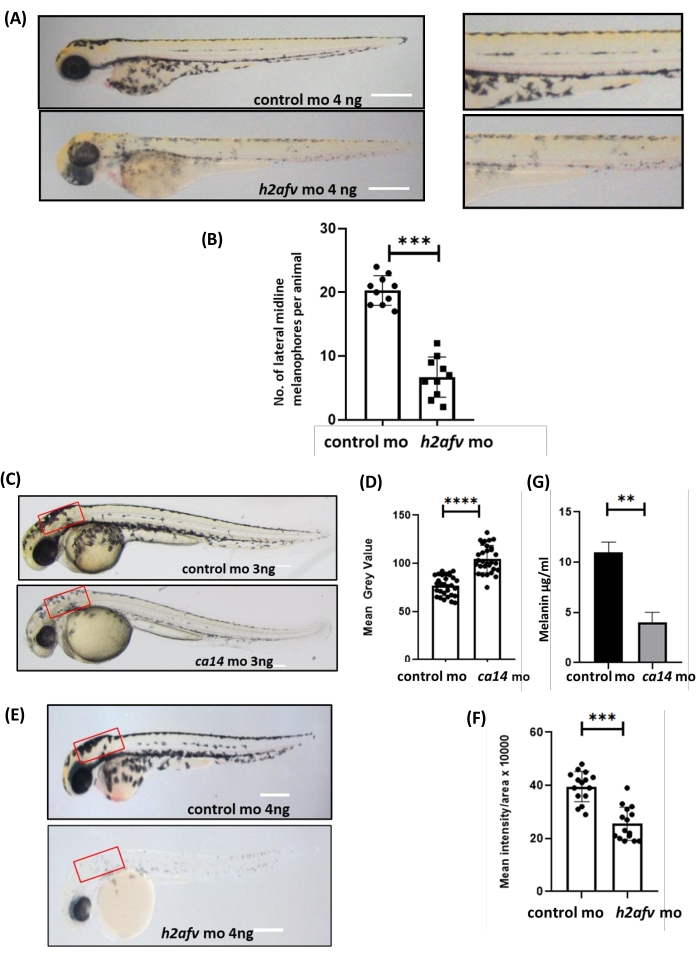
Figure 2: Counting lateral midline melanophores, measuring mean gray value, and melanin content assay. Brightfield microscopic images of the lateral view of morphants were used to quantify melanin content using ImageJ software. (A) Control and Histone variant h2afv (h2afv) morphants at 3 dpf; on the right are zoomed-in trunk regions of these embryos depicting melanocyte numbers. (B) The number of melanophores in h2afv morphants is drastically reduced relative to control, n > 10 each in 3 biological replicates. (C) Representative image of control vs carbonic anhydrase 14 morphants at 2 dpf. (D) Melanin quantification of ca14 vs control morphants, n > 10 across 3 biological replicates. (E) Lateral view of h2afv vs control morphants at 2 dpf. (F) Melanin content quantification of h2afv vs control morphants. Red rectangles represent ROI chosen for ImageJ-based analysis. (G) Melanin content assay performed on morpholino-injected embryos for quantifying the total melanin content. Mean of three independent experiments ± SEM.; **P < 0.01, **** P < 0.0001. Student's t-test; error bars are mean ± standard error of the mean (SEM). Scale bars = 100 µm. Abbreviations: MO = morpholino; dpf = days post fertilization; ROI = region of interest. Please click here to view a larger version of this figure.
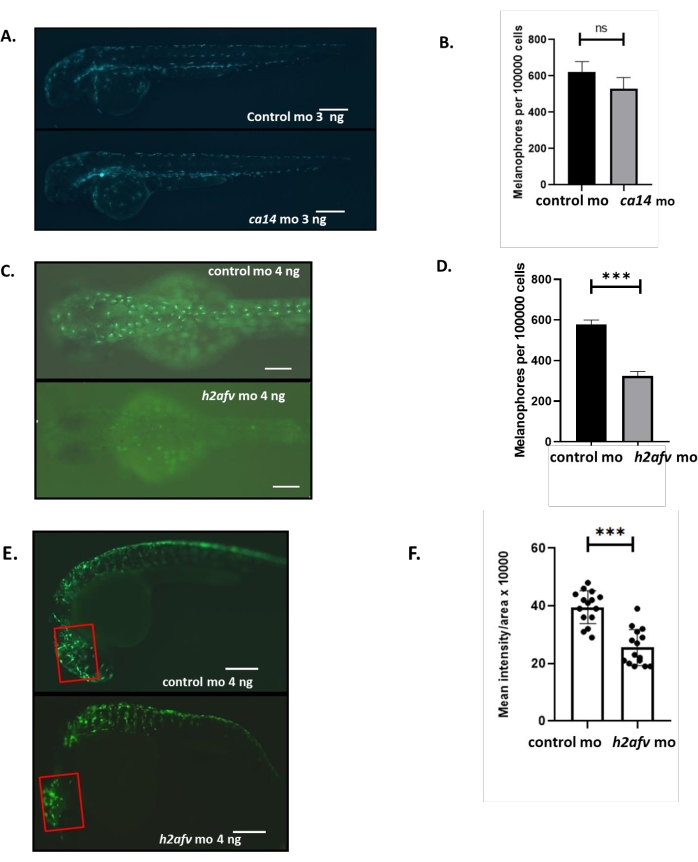
Figure 3: Quantifying fluorescently labeled cells using FACS and measuring mean fluorescence intensity. (A) Fluorescence images of ca14 and control morphant embryos at 2 dpf from Tg (ftyrp1: GFP) line, where the differentiating melanocytes are marked by GFP expression. (B) Number of melanophores in ca14 and control morphant embryos at 2 dpf remain unchanged, n = 3 biological replicates. (C) Fluorescence images at 2 dpf of h2afv and control morphants from Tg(ftyrp1:GFP) line. (D) The number of melanophores in h2afv morphants is significantly reduced compared to the control morphant embryos at 2 dpf, n = 3 biological replicates. (E) Fluorescence images of control and h2afv morphant embryos at 36 hpf from Tg(mitfa:GFP) with labeled melanocytes. (F) MFI per animal is calculated using ImageJ software and represented as a bar graph depicting individual data sets. MFI is significantly reduced in h2afv morphants compared to control. Red rectangle represents ROI chosen for ImageJ software-based analysis. n = 3 biological replicates; ***P < 0.001, ****P < 0.0001. Student's t-test; error bars are mean ± standard error of the mean (SEM); scale bars = 100 µm. Abbreviations: MO = morpholino; FACS = fluorescence-activated cell sorting; GFP = green fluorescent protein; dpf = days post fertilization; ROI = region of interest; MFI = mean fluorescence intensity. Please click here to view a larger version of this figure.
Supplemental Figure S1: Quantitation of head melanophores. (A) Dorsal view of h2afv and control morphants at 48 hpf showing head melanophores. Red box represents the ROI. Keeping the area of interest constant, the number of head melanophores can be quantified manually. (B) Number of head melanophores significantly reduced upon h2afv knockdown. Bars represent geometric mean with 95% CI of the number of head melanophores per embryo with >30 embryos each. Scale bars = 100 µm. Abbreviations: MO = morpholino; CI = confidence interval; hpf = hours post fertilization; ROI = region of interest. Please click here to download this File.
Supplemental Figure S2: Microarray of human primary melanocytes treated with PTU. Heat map shows the expression of pigmentation genes (mitf, tyrosinase, dct, mc1r) upon treatment in human primary melanocytes. Abbreviations: PTU = 1-phenyl-2-thiourea; tyr = tyrosinase. Please click here to download this File.
