Strongyloides stercoralis har lenge blitt oversett som et viktig menneskelig patogen sammenlignet med de mer anerkjente krokormene og rundorm Ascaris lumbricoides1. Tidligere studier av ormbyrde undervurderte ofte utbredelsen av S. stercoralis på grunn av den lave følsomheten til vanlige diagnostiske metoder for S. stercoralis2. De siste årene har epidemiologiske studier basert på forbedrede diagnostiske verktøy anslått at den sanne forekomsten av S. stercoralis-infeksjoner er mye høyere enn tidligere rapportert, omtrent 610 millioner mennesker over hele verden2.
Både S. stercoralis og andre Strongyloides-arter, inkludert den nært beslektede rotteparasitten og den vanlige laboratoriemodellen S. ratti, har en uvanlig livssyklus som er fordelaktig for eksperimentelle genomiske studier fordi den består av både parasittiske og frittlevende (miljømessige) generasjoner3 (figur 1). Spesielt kan både S. stercoralis og S. ratti sykle gjennom en enkelt frittlevende generasjon. Den frittlevende generasjonen består av post-parasittiske larver som utvikler seg til frittlevende voksne menn og kvinner; alle avkom av de frittlevende voksne utvikler seg til smittende larver, som må infisere en vert for å fortsette livssyklusen. Videre kan denne miljømessige eller frittlevende generasjonen eksperimentelt manipuleres i laboratoriet. Fordi frittlevende Strongyloides voksne og C. elegans voksne deler lignende morfologi, kan teknikker som intragonadal mikroinjeksjon som opprinnelig ble utviklet for C. elegans tilpasses for bruk med frittlevende voksen Strongyloides 4,5. Mens DNA generelt introduseres til frittlevende voksne kvinner, kan både menn og kvinner av Strongyloides mikroinjiseres6. Dermed er funksjonelle genomiske verktøy tilgjengelige for å forhøre mange aspekter av Strongyloides biologi. Andre parasittiske nematoder mangler en frittlevende generasjon, og som et resultat er det ikke så lett egnet til funksjonelle genomiske teknikker3.
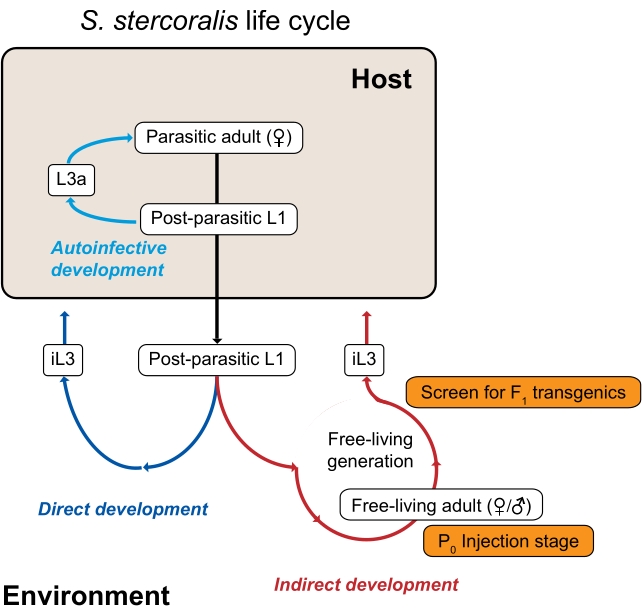
Figur 1: Strongyloides stercoralis livssyklus. S. stercoralis parasittiske kvinner bor i tynntarmen til sine pattedyrverter (mennesker, ikke-menneskelige primater, hunder). De parasittiske hunnene reproduserer ved parthenogenese og legger egg i tynntarmen. Eggene klekkes mens de fortsatt er inne i verten i postparasittiske larver, som deretter overføres inn i miljøet med avføring. Hvis de postparasittiske larver er mannlige, utvikler de seg til frittlevende voksne menn. Hvis de postparasittiske larver er kvinnelige, kan de enten utvikle seg til frittlevende voksne kvinner (indirekte utvikling) eller tredjetrinns infektive larver (iL3s; direkte utvikling). De frittlevende hannene og hunnene reproduserer seksuelt for å skape avkom som er begrenset til å bli iL3s. Under visse forhold kan S. stercoralis også gjennomgå autoinfeksjon, der noen av de postparasittiske larver forblir inne i vertstarmen i stedet for å passere inn i miljøet i avføring. Disse larvene kan utvikle seg til autoinfektive larver (L3a) inne i verten, trenge gjennom tarmveggen, migrere gjennom kroppen og til slutt gå tilbake til tarmen for å bli reproduktive voksne. Livssyklusen til S. ratti er lik, bortsett fra at S. ratti infiserer rotter og ikke har en autoinfective syklus. Miljøgenerasjonen er nøkkelen til å bruke Strongyloides arter til genetiske studier. De frittlevende voksne hunnene (P0) kan mikroinjiseres; deres avkom, som alle vil bli iL3s, er potensielle F1 transgenics. Denne figuren er endret fra Castelletto et al. 3. Klikk her for å se en større versjon av denne figuren.
S. stercoralis deler mange aspekter av sin biologi med andre gastrointestinale menneske-parasittiske nematoder, inkludert vertsinvasjon og vertsimmunmodulering. For eksempel smitter human-parasittiske krokorm i slekten Necator og Ancylostoma også ved hudpenetrasjon, navigerer på samme måte gjennom kroppen, og til slutt bor som parasittiske voksne i tynntarmen7. Dermed bruker mange gastrointestinale nematoder sannsynligvis vanlig sensorisk oppførsel og immununndragelsesteknikker. Som et resultat vil kunnskapen fra Strongyloides utfylle funn i andre mindre genetisk gjennomførbare nematoder og føre til en mer fullstendig forståelse av disse komplekse og viktige parasittene.
Denne mikroinjeksjonsprotokollen skisserer metoden for å introdusere DNA i Strongyloides frittlevende voksne kvinner for å lage transgene og mutante avkom. Belastningsvedlikeholdskravene, inkludert utviklingstid for voksne ormer for mikroinjeksjoner og innsamling av transgent avkom, er beskrevet. Protokoller og en demonstrasjon av den komplette mikroinjeksjonsteknikken, sammen med protokoller for dyrking og screening av transgent avkom, er inkludert, sammen med en liste over alt nødvendig utstyr og forbruksvarer.
