Virusliknande partiklar (VLPs) liknar den inhemska viruspartikelstrukturen samtidigt som de saknar det virala genomet, vilket ger en hög säkerhetsprofil1,2. VLP representerar en individuell klass av vacciner som utvecklas alltmer på grund av deras höga immunogenicitet3,4,5,6,7. Detta gäller särskilt membran-hölje VLPs, vilket möjliggör visning av inte bara homologa virala ytan antigener men också heterologous antigener såsom tumör antigener8,9,10. Figur 1 ger en föredömlig översikt över strukturen hos en omsluten antigendekorerad VLP. Under utvecklingsprocessen av VLP-baserade vacciner är analyser oumbärliga för att möjliggöra analys av respektive målantigen som visas på VLP-ytan. Sådana analyser bör vara avgörande för att klargöra sammansättningen av ett partikelvaccin: i) Är VLP dekorerade med respektive ytantigen? ii) Har ytantigenet behållit sin ursprungliga struktur, vilket framgår av epitopigenkänning av neutraliserande antikroppar (bNAbs) och iii) kan VLP:s strukturella integritet bekräftas på grund av påvisande av det virala protein som förmedlar VLP-bildandet?
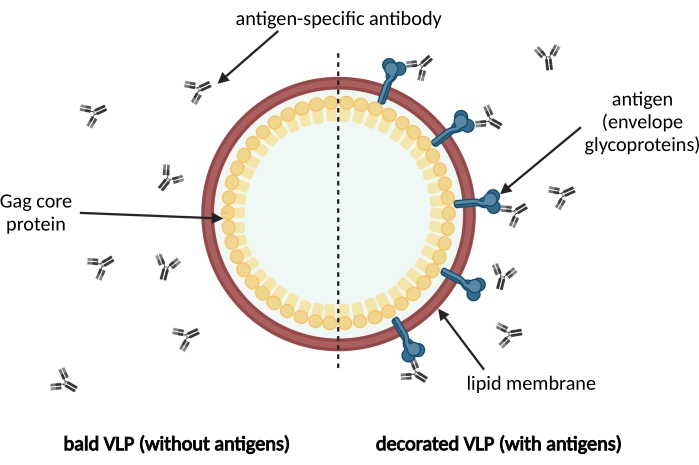
Bild 1: Schematisk illustration av en membransluten VLP. VLPs bildas av omogna prekursor Gag kärnproteiner och omges av ett lipidmembran som härrör från värdcellen. Antigenerna, t.ex. kuvertglykoproteiner, införlivas i lipidmembranet och visas på VLP:s yta (till höger). Antigenspecifika antikroppar känner igen antigenet. Till vänster visas en skallig VLP utan antigendekoration. Klicka här för att se en större version av den här figuren.
Särskilt VLPs bildas av viral gruppspecifik antigen (Gag) core precursor protein p55 av humant immunbristvirus typ 1 (HIV-1) är föredragna byggnadsställningar för antigen display i vaccin utveckling som många antikroppar, och ELISA kit finns tillgängliga, vilket möjliggör kvantifiering av dessa VLPs11,12. Hiv-1-kuvertet glykoproteiner (Env), nämligen transmembranproteinet gp41 (gp41-TM) och den lösliga ytanheten gp120 (gp120-SU) som bildar heterodimerer, införlivas i membranhöljet av partiklar och är viktiga målantigener för utveckling av vacciner mot HIV-infektion13,14,15 . Visning av neutraliseringskänsliga epitoper i dessa målantigener är en förutsättning för att framkalla ett i stort sett neutraliserande antikroppssvar i vacciner. Förutom ett T-cellssvar riktat mot Gag-proteinerna anses detta vara en viktig korrelat av skydd mot HIV-infektion16. Följaktligen, och vid utformning och produktion av VLP dekorerade med målantigenkandidater, utgör den efterföljande analysen av kvaliteten på de visade antigenerna ett kritiskt steg i processen för vaccinutveckling.
Immunoprecipitation (IP) är en allmänt använd teknik för detektion av protein-proteininteraktioner och rening av proteinkomplex i liten skala17. Barret et al. först rapporterades om utvecklingen av immateriella rättigheter 1960, men denna metod har ständigt förbättrats ytterligare. IP möjliggör avskiljning och isolering av ett målantigen (byte) från en lösning genom att använda en antigenspecifik antikropp (bete) immobiliserad genom koppling till pärlor18,19. I detta protokoll visar vi en variation av den klassiska IP-applikationen med membran-enveloped p55 Gag-formade VLPs som byte och bNAbs som känner igen neutraliserande känsliga epitoper i kuvertproteinerna som visas på ytan av VLPs som beteproteiner. Den framgångsrika tillämpningen av denna VLP-fångstanalys underlättar förutsägelsen om huruvida de testade antigenpositiva VLPs kommer att kunna framkalla en neutraliserande B-cellsrespons hos vaccinerade personer. Sådana immunogenica egenskaper hos VLP-baserade vaccinkandidater demonstreras ofta i smådjursmodeller20,21,22.
För att bedöma kvaliteten på den nyutvecklade VLP-vaccinkandidaten har VLP-insamlingsanalyser framgångsrikt använts5,23,24. Antalet publicerade metoder är dock begränsat. VLP fångst analys som presenteras här börjar med immobilisering av Env-specifika bNAbs på protein G-konjugerade pärlor, som binder till Fc regionen av däggdjur-härledda antikroppar. Typiska matriser för immobilisering av den antikropp som valts är agaros eller magnetiska pärlor. Magnetiska pärlor är dock gynnsamma för applikationer med hög genomströmning25. I nästa steg fångas VLP:er som visar målantigenet av bNAb-belagda pärlor. De bildade immunkomplexen bestående av Env-positiva VLPs och immobiliserade bNAbs berikas lätt med hjälp av en magnet. De isolerade immunkomplexen eluteras i det sista steget. Därefter kan VLPs biokemiskt karakteriseras. Här utförde vi western blot-analys med p55 Gag viral kärna protein-specifika antikroppar för att visa att det utfällda målet Env antigener inte bara hyser neutraliserande känsliga epitopes men visades också på Gag-bildade VLPs. Dessutom ökar upptäckten av viruskärnan Gag-proteiner känsligheten hos fångstanalysen eftersom Gag-proteiner är mer rikliga än Env i en VLP. I HIV-1 finns Env-proteiner endast på ett ensiffrigt eller tvåsiffrigt nummer26, medan mer än 3 500 gagmolekyler utgör kärnan i en partikel27.
Jämfört med andra tekniker för undersökning av interaktioner mellan protein och protein28,29 ger VLP-insamlingsanalysen en alternativ metod för forskningslaboratorier som inte har tillgång till dyra analysinstrument. Till exempel kan transmissionselektronmikroskopisk analys (TEM), ytplasmonresonansspektroskopi (SPR) och nanopartikelspårningsanalys (NTA) vara kostnadsintensiva. Den avskiljningsanalys som presenteras här tillåter också senare subjektering av fångade antigenpositiva VLP-prover till ytterligare proteinkarakterisering, t.ex. med hjälp av gelelektrofores, immunoblotting, elektronmikroskopi respektive masspektrometri (MS). Med tanke på att målantigenets ursprungliga struktur bevaras under VLP-fångstanalysen kan även prestandan hos en inbyggd SIDA och efterföljande immunblåsningstekniker användas.
VLP-fångstanalysen representerar en lättanvänd och känslig metod för att undersöka dekorationen av VLPs med målantigener som exponerar neutraliserande känsliga epitoper, och därmed deras användbarhet som framtida vaccinkandidater.
