Stoffwechsel ist der zelluläre Prozess der Energieerzeugung. Der Zellstoffwechsel umfasst mehrere Wege, einschließlich Glykolyse, oxidative Phosphorylierung und Glutaminolyse. Gesunde Zellen nutzen diese Stoffwechselwege, um Energie für die Proliferation und Funktion zu erzeugen, wie zum Beispiel die Produktion von Zytokinen durch Immunzellen. Viele Krankheiten, einschließlich Stoffwechselstörungen, Krebs und Neurodegeneration, sind durch einen veränderten Zellstoffwechsel gekennzeichnet1. Zum Beispiel haben einige Krebszelltypen erhöhte Glykolyseraten, selbst in Gegenwart von Sauerstoff, um Moleküle für die Synthese von Nukleinsäuren, Proteinen und Lipiden zu erzeugen2,3. Dieses Phänomen, das als Warburg-Effekt bekannt ist, ist ein Kennzeichen vieler Krebsarten, einschließlich Brustkrebs, Lungenkrebs und Glioblastome4. Aufgrund der Veränderungen des Zellstoffwechsels, die mit dem Fortschreiten der Krebserkrankung verbunden sind, kann der Zellstoffwechsel ein Surrogat-Biomarker für die Arzneimittelreaktion sein5,6. Darüber hinaus ist das Verständnis der Arzneimittelwirksamkeit auf zellulärer Ebene von entscheidender Bedeutung, da die Zellheterogenität bei Individuen zu unterschiedlichen Arzneimittelreaktionen führen kann7,8.
Technologien, die Veränderungen im Zellstoffwechsel identifizieren und quantifizieren, sind für Studien zu Krebs und Medikamentenreaktion unerlässlich. Chemische und Proteinanalysen werden verwendet, um den Stoffwechsel von Zellen oder Geweben zu bewerten, aber es fehlen Einzelzellauflösungen und räumliche Informationen. Metabolische Plattenleser-basierte Assays können den pH- und Sauerstoffverbrauch in der Probe im Laufe der Zeit und die anschließende metabolische Störung durch Chemikalien messen. Über den pH-Wert kann die extrazelluläre Versauerungsrate (ECAR) berechnet werden, die einen Einblick in die glykolytische Aktivität der Zellen gibt9. Bildgebende Ganzkörpermethoden, einschließlich 2-[Fluor-18] Fluor-D-Glucose-Positronen-Emissions-Tomographie (FDG PET) und Magnetresonanzspektroskopie (MRS), sind nichtinvasive Bildgebungsmodalitäten, die klinisch verwendet werden, um Tumorrezidive und Arzneimittelwirksamkeit durch metabolische Messungen zu identifizieren10,11,12,13,14.
FDG-PET bildet die Gewebeaufnahme von FDG, einem radioaktiv markierten Glukoseanalogon, ab. Die erhöhte Aufnahme von FDG-PET durch Tumore im Verhältnis zum umgebenden Gewebe ist auf den Warburg-Effekt zurückzuführen12,13. MRS bildet gemeinsame Kerne von Molekülen ab, die für den Stoffwechsel verwendet werden, wie 13C und 31P, und kann dynamische Informationen darüber erhalten, wie sich der Stoffwechsel als Reaktion auf Reize wie Bewegung oder Essen verändert14. Obwohl FDG-PET und MRS klinisch eingesetzt werden können, fehlt diesen Technologien die räumliche Auflösung, um die intratumorale Heterogenität aufzulösen. Ebenso werden Sauerstoffverbrauchsmessungen an einer Massenpopulation von Zellen durchgeführt. Die Autofluoreszenzbildgebung überwindet das räumliche Auflösungshindernis dieser Technologien und bietet eine nichtinvasive Methode zur Quantifizierung des Zellstoffwechsels.
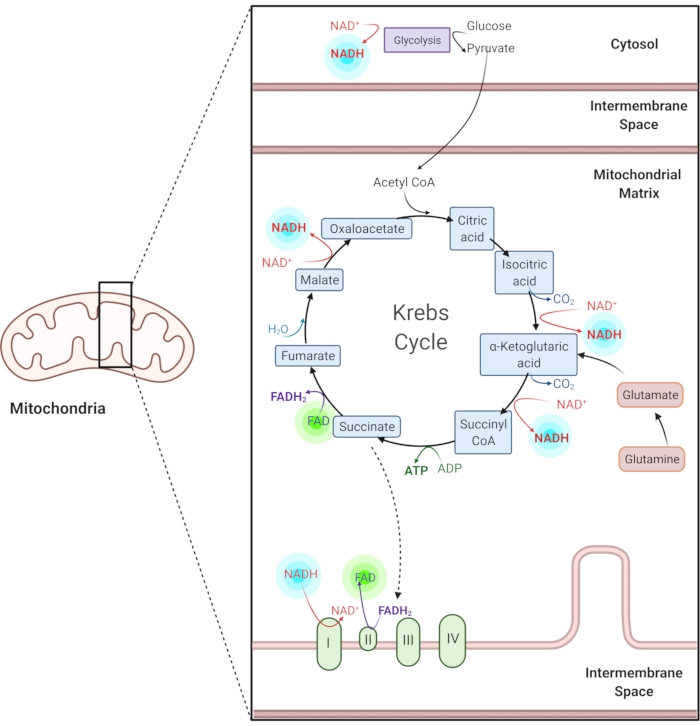
Abbildung 1: NADH und FAD in gemeinsamen Stoffwechselwegen. NADH und FAD sind Coenzyme, die in der Glykolyse, im Krebszyklus und in der Elektronentransportkette verwendet werden. Die Autofluoreszenzbildgebung dieser Moleküle liefert Informationen über den Zellstoffwechsel. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
Reduziertes Nicotinamidadenin (Phosphat) Dinukleotid (NAD(P)H) und oxidiertes Flavinadenindinukleotid (FAD) sind Coenzyme von Stoffwechselreaktionen, einschließlich Glykolyse, oxidativer Phosphorylierung und Glutaminolyse (Abbildung 1). Sowohl NAD(P)H als auch FAD sind autofluoreszierend und bieten einen endogenen Kontrast für die Fluoreszenzbildgebung1,15. NADPH hat ähnliche fluoreszierende Eigenschaften wie NADH. Aus diesem Grund wird NAD(P)H häufig verwendet, um das kombinierte Signal von NADH und NADPH2,16 darzustellen.
Die Fluoreszenzlebensdauerbildgebung (FLIM) quantifiziert die Fluoreszenzlebensdauer oder die Zeit, für die sich ein Fluorophor im angeregten Zustand befindet. Fluoreszenzlebensdauern reagieren auf die Mikroumgebung der Fluorophore und liefern Informationen über den Zellstoffwechsel17. NAD(P)H und FAD können in Zellen entweder in proteingebundenen oder freien Konformationen existieren, von denen jede eine andere Lebensdauer hat. Freies NAD(P)H hat eine kürzere Lebensdauer als proteingebundenes NAD(P)H; Umgekehrt hat die freie FAD eine längere Lebensdauer als die gebundenen FAD18,19. Die Lebensdauern und Lebensdauerkomponentengewichte können aus Fluoreszenz-Lebensdauerzerfallsdaten durch Gl. (1)20 quantifiziert werden:
I(t) = α 1e-t/τ1 + α 2e-t/τ2 + C (1)
Eq (1) stellt die normierte Fluoreszenzintensität als Funktion der Zeit dar. Die α 1 und α 2 in dieser Gleichung stellen die proportionalen Komponenten der kurzen und langen Lebensdauer (α 1+ α 2=1) dar, τ1 und τ2 die kurze bzw. lange Lebensdauer und C das Hintergrundlicht7,20. Die amplitudengewichtete Lebensdauer, hier dargestellt als τm, wird mit Gleichung (2) berechnet.
τm= α 1τ1+ α 2τ2 (2)
Eine mittlere Lebensdauer kann durch Mittelung von “t” über den Intensitätszerfall des Fluorophors berechnet werden, was für einen zweiexponentiellen Zerfall durch Gl. (3)17,21 dargestellt wird.
τ*m= (α 1τ12+ α 2τ22)/ (α 1τ1+ α 2τ2) (3)
Ein Fluoreszenzintensitätsbild kann aus dem Lebensdauerbild berechnet werden, indem der Fluoreszenzlebensdauerzerfall integriert wird. Die Autofluoreszenzbildgebung ist eine zerstörungsfreie und markierungsfreie Methode, mit der der Stoffwechsel lebender Zellen mit subzellulärer Auflösung charakterisiert werden kann. Das optische Redoxverhältnis liefert eine optische Analogmetrik des chemischen Redoxzustands der Zelle und wird als Verhältnis von NAD(P)H- und FAD-Intensitäten berechnet. Obwohl die Formel zur Berechnung des optischen Redoxverhältnisses nicht standardisiert ist22,23,24,25, wird sie hier als die Intensität von FAD über die kombinierten Intensitäten von NAD(P)H und FAD definiert. Diese Definition wird verwendet, weil die summierte Intensität im Nenner die Metrik zwischen 0 und 1 normalisiert und das erwartete Ergebnis der Cyanidhemmung eine Abnahme des Redoxverhältnisses ist. Die Fluoreszenzlebensdauern von freiem NAD(P)H und FAD liefern Einblicke in Veränderungen in der metabolischen Lösungsmittelmikroumgebung, einschließlich pH-Wert, Temperatur, Nähe zu Sauerstoff und Osmolarität17.
Veränderungen in der Fluoreszenzlebensdauer der gebundenen Fraktionen von NAD(P)H und FAD können auf die Nutzung des Stoffwechselwegs und den substratspezifischen Stoffwechsel hinweisen26. Komponentengewichte können für Änderungen der freien zur gebundenen Fraktion der Coenzyme interpretiert werden18,19. Insgesamt ermöglichen diese quantitativen Autofluoreszenz-Lebenszeitmetriken die Analyse des Zellstoffwechsels, und die Autofluoreszenz-Bildgebung wurde zur Identifizierung von Neoplasmen aus normalem Gewebe verwendet27,28, zur Charakterisierung von Stammzellen29,30, zur Bewertung der Immunzellfunktion31,32,33,34,35, zur Messung der neurologischen Aktivität36, 37,38, und Verständnis der Arzneimittelwirksamkeit bei Krebsarten wie Brustkrebs und Kopf-Hals-Krebs21,39,40,41,42. Hochauflösende Autofluoreszenzbildgebung kann mit Bildsegmentierung für die Einzelzellanalyse und Quantifizierung der intrapopulationären Heterogenität kombiniert werden43,44,45,46,47.
NAD(P)H und FAD können auf Einzelphotonen- oder Multiphotonen-Fluoreszenzmikroskopen abgebildet werden, die für die Intensitäts- oder Lebensdauerbildgebung konfiguriert sind. Bei Einzelphotonenmikroskopen werden NAD(P)H und FAD aufgrund üblicher Laserquellen bei diesen Wellenlängen typischerweise bei Wellenlängen von 375-405 nm bzw. 488 nm angeregt48. Bei der Zwei-Photonen-Fluoreszenzanregung werden NAD(P)H und FAD bei Wellenlängen von etwa 700 bis 750 nm bzw. 700 bis 900 nm bzw. 15,49 anregen. Sobald die Fluorophore angeregt sind, emittieren NAD(P)H und FAD Photonen bei Wellenlängen zwischen ~410 nm bis ~490 nm bzw. ~510 nm bis ~640 nm15. Die maximalen Emissionswellenlängen NAD(P)H und FAD betragen etwa 450 nm bzw. 535 nm48.
Aufgrund ihrer unterschiedlichen Anregungs- und Emissionswellenlängen kann die Fluoreszenz der beiden metabolischen Coenzyme spektral isoliert werden. Ein Verständnis der spektralen Eigenschaften von NAD(P)H und FAD ist für das Design und die Optimierung von Autofluoreszenz-Bildgebungsprotokollen notwendig. Cyanid ist ein Elektrontransportkette (ETC) -Komplex-IV-Inhibitor. Die Auswirkungen von Cyanid auf den Zellstoffwechsel und die Autofluoreszenzintensitäten und Lebensdauern von NAD(P)H und FAD in Zellen sind gut charakterisiert27,40. Daher ist ein Cyanid-Störungsexperiment ein wirksames Mittel zur Validierung von NAD(P)H- und FAD-Bildgebungsprotokollen. Ein erfolgreiches Cyanidexperiment gibt die Gewissheit, dass das NAD(P)H- und FAD-Bildgebungsprotokoll verwendet werden kann, um den Stoffwechsel unbekannter Gruppen oder Störungen zu beurteilen.
