Metabolisme er den cellulære prosessen med å produsere energi. Cellulær metabolisme omfatter flere veier, inkludert glykolyse, oksidativ fosforylering og glutaminolyse. Sunn celler bruker disse metabolske veiene for å generere energi til spredning og funksjon, for eksempel produksjon av cytokiner av immunceller. Mange sykdommer, inkludert metabolske forstyrrelser, kreft og nevrodegenerasjon, er preget av endret cellulær metabolisme1. For eksempel har noen kreftcelletyper forhøyede glykolysehastigheter, selv i nærvær av oksygen, for å generere molekyler for syntese av nukleinsyrer, proteiner og lipider2,3. Dette fenomenet, kjent som Warburg-effekten, er et kjennetegn på mange krefttyper, inkludert brystkreft, lungekreft og glioblastomer4. På grunn av endringene i cellulær metabolisme forbundet med kreftprogresjon, kan cellulær metabolisme være en surrogatbiomarkør for legemiddelrespons5,6. Videre er det avgjørende å forstå legemiddeleffekten på cellenivå, da celle heterogenitet kan føre til forskjellige legemiddelresponser hos personer7,8.
Teknologier som identifiserer og kvantifiserer endringer i cellulær metabolisme er avgjørende for studier av kreft og legemiddelrespons. Kjemiske og proteinanalyser brukes til å evaluere metabolismen av celler eller vev, men mangler encellet oppløsning og romlig informasjon. Metabolske plateleserbaserte analyser kan måle pH- og oksygenforbruk i prøven over tid og den påfølgende metabolske perturbasjonen av kjemikalier. pH kan brukes til å beregne den ekstracellulære forsuringshastigheten (ECAR), som gir et innblikk i cellenes glykolytiske aktivitet9. Avbildningsmetoder for hele kroppen, inkludert 2-[fluor-18] fluor-D-glukose positronutslippstomografi (FDG PET) og magnetisk resonansspektroskopi (MRS), er ikke-invasive avbildningsmodaliteter som brukes klinisk for å identifisere tumor gjentakelse og legemiddeleffekt gjennom metabolske målinger10,11,12,13,14.
FDG-PET tar bilder av vevsopptaket til FDG, en radiomerket glukoseanalog. Økt opptak av FDG-PET av svulster i forhold til omkringliggende vev skyldes Warburg-effekten12,13. MRS-bilder vanlige kjerner av molekyler som brukes til metabolisme, for eksempel 13C og 31P, og kan få dynamisk informasjon om hvordan metabolisme endres som respons på stimuli, for eksempel trening eller spising14. Selv om FDG-PET og MRS kan brukes klinisk, mangler disse teknologiene den romlige oppløsningen for å løse intratumoral heterogenitet. På samme måte gjøres oksygenforbruksmålinger på en bulkpopulasjon av celler. Autofluorescence imaging overvinner den romlige oppløsningen hinderet for disse teknologiene og gir en ikke-invasiv metode for kvantifisering av cellulær metabolisme.
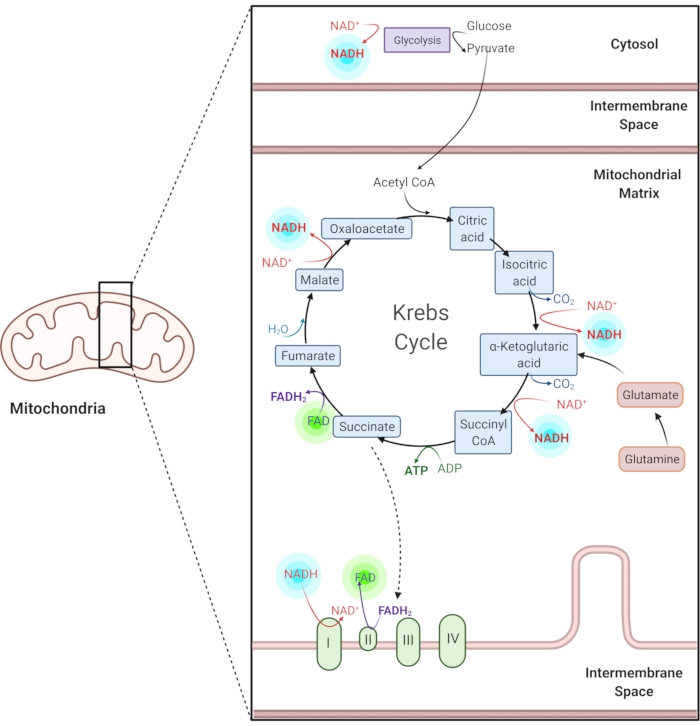
Figur 1: NADH og FAD i vanlige metabolske veier. NADH og FAD er koenzymer som brukes i glykolyse, Krebs-syklusen og elektrontransportkjeden. Autofluorescence avbildning av disse molekylene gir informasjon om cellulær metabolisme. Klikk her for å se en større versjon av denne figuren.
Redusert nikotinamid adenin (fosfat) dinukleotid (NAD(P)H) og oksidert flavin adenin dinukleotid (FAD) er koenzymer av metabolske reaksjoner, inkludert glykolyse, oksidativ fosforylering og glutaminolyse (figur 1). Både NAD(P)H og FAD er autofluorescerende og gir endogen kontrast for fluorescensavbildning1,15. NADPH har lignende fluorescerende egenskaper som NADH. På grunn av dette brukes NAD(P)H ofte til å representere det kombinerte signalet til NADH og NADPH2,16.
Fluorescens livstidsavbildning (FLIM) kvantifiserer fluorescenslevetiden eller tiden som en fluorofor er i spent tilstand. Fluorescenslevetidene reagerer på mikromiljøet i fluoroforene og gir informasjon om cellulær metabolisme17. NAD(P)H og FAD kan eksistere i celler i enten proteinbundne eller frie konformasjoner, som hver har forskjellig levetid. Gratis NAD(P)H har kortere levetid enn proteinbundet NAD(P)H; på den annen side har fri FAD lengre levetid enn bundet FAD18,19. Levetids- og levetidskomponentvektene kan kvantifiseres fra fluorescenslevetidsforfallsdata gjennom Eq. (1)20:
I(t) = α 1e-t/τ1 + α 2e-t/τ2 + C (1)
Eq (1) representerer den normaliserte fluorescensintensiteten som en tidsfunksjon. Den α 1 og α 2 i denne ligningen representerer de proporsjonale komponentene i kort og lang levetid (henholdsvis α 1+ α 2 = 1), henholdsvis τ1 og τ2 representerer henholdsvis korte og lange levetider, og C står for bakgrunnslys7,20. Den amplitudevektede levetiden, representert her som τm, beregnes ved hjelp av Eq. (2).
τm= α 1τ1+ α 2τ2 (2)
En gjennomsnittlig levetid kan beregnes ved å beregne “t” over fluoroforens intensitetsforfall, som for et to eksponentielt forfall er vist av Eq. (3)17,21.
τ*m= (α 1τ12+ α 2τ22)/ (α 1τ1+ α 2τ2) (3)
Et fluorescensintensitetsbilde kan beregnes fra levetidsbildet ved å integrere fluorescenslevetidens forfall. Autofluorescence avbildning er en ikke-ødeleggende og etikettfri metode som kan brukes til å karakterisere metabolismen av levende celler med en subcellulær oppløsning. Det optiske redoksforholdet gir en optisk analog beregning av cellens kjemiske redokstilstand og beregnes som forholdet mellom NAD(P)H- og FAD-intensiteter. Selv om formelen for beregning av det optiske redoksforholdet ikke er standardisert22,23,24,25, er det her definert som intensiteten av FAD over de kombinerte intensitetene til NAD (P)H og FAD. Denne definisjonen brukes fordi den summerte intensiteten i nevneren normaliserer metrikkverdien mellom 0 og 1, og det forventede resultatet av cyanidinhiberingen er en reduksjon i redoksforholdet. Fluorescenslevetidene til fri NAD(P)H og FAD gir innsikt i endringer i det metabolske løsningsmiddelmikromiljøet, inkludert pH, temperatur, nærhet til oksygen og osmolaritet17.
Endringer i fluorescenslevetiden for de bundne fraksjonene av NAD(P)H og FAD kan indikere metabolsk baneutnyttelse og substratspesifikk metabolisme26. Komponentvekter kan tolkes for endringer i fri til den bundne brøkdelen av koenzymene18,19. Til sammen tillater disse kvantitative autofluorescence levetidsmålingene analysen av cellulær metabolisme, og autofluorescence imaging har blitt brukt til å identifisere neoplasmer fra normale vev27,28, karakterisere stamceller29,30, evaluere immuncellefunksjon31,32,33,34,35, måle nevrologisk aktivitet36, 37,38, og forstå legemiddeleffekt i krefttyper som brystkreft og hode- og nakkekreft21,39,40,41,42. Høyoppløselig autofluorescence-avbildning kan kombineres med bildesegmentering for encellet analyse og kvantifisering av intrapopulerings heterogenitet43,44,45,46,47.
NAD(P)H og FAD kan avbildes på enkeltfoton- eller multifotonfluorescensmikroskoper som er konfigurert for intensitet eller livstidsavbildning. For enkeltfotonmikroskoper er NAD(P)H og FAD vanligvis begeistret for bølgelengder på henholdsvis 375-405 nm og 488 nm på grunn av vanlige laserkilder ved disse bølgelengdene48. I to-foton fluorescens excitation, NAD (P)H og FAD vil begeistre ved bølgelengder på ca 700 til 750 nm og 700 til 900 nm, henholdsvis15,49. Når fluoroforene er begeistret, avgir NAD(P)H og FAD fotoner ved bølgelengder mellom ~ 410 nm til ~ 490 nm og ~ 510 nm til ~ 640 nm, henholdsvis15. NAD(P)H og FAD maxima utslipp bølgelengder er ca 450 nm og 535 nm, henholdsvis48.
På grunn av deres forskjellige eksitasjons- og utslippsbølgelengder, kan fluorescensen til de to metabolske koenzymene isoleres spektralt. En forståelse av spektralkarakteristikkene til NAD(P)H og FAD er nødvendig for design og optimalisering av autofluorescence imaging protokoller. Cyanid er en elektrontransportkjede (ETC) kompleks IV-hemmer. Effektene av cyanid på cellulær metabolisme og autofluorescensintensiteter og levetider for NAD(P)H og FAD i celler er godt karakterisert27,40. Derfor er et cyanidperturbasjonseksperiment et effektivt middel for å validere NAD(P)H- og FAD-bildebehandlingsprotokoller. Et vellykket cyanideksperiment gir tillit til at NAD(P)H- og FAD-bildebehandlingsprotokollen kan brukes til å vurdere metabolismen av ukjente grupper eller perturbasjoner.
