El metabolismo es el proceso celular de producción de energía. El metabolismo celular abarca múltiples vías, incluyendo la glucólisis, la fosforilación oxidativa y la glutaminólisis. Las células sanas utilizan estas vías metabólicas para generar energía para la proliferación y la función, como la producción de citoquinas por las células inmunes. Muchas enfermedades, incluyendo trastornos metabólicos, cáncer y neurodegeneración, se caracterizan por un metabolismo celular alterado1. Por ejemplo, algunos tipos de células cancerosas tienen tasas elevadas de glucólisis, incluso en presencia de oxígeno, para generar moléculas para la síntesis de ácidos nucleicos, proteínas y lípidos2,3. Este fenómeno, conocido como el efecto Warburg, es un sello distintivo de muchos tipos de cáncer, incluyendo el cáncer de mama, el cáncer de pulmón y los glioblastomas4. Debido a las alteraciones del metabolismo celular asociadas a la progresión del cáncer, el metabolismo celular puede ser un biomarcador sustituto de la respuesta farmacológica5,6. Además, la comprensión de la eficacia del fármaco a nivel celular es crucial, ya que la heterogeneidad celular puede conducir a diferentes respuestas farmacológicas en los individuos7,8.
Las tecnologías que identifican y cuantifican los cambios en el metabolismo celular son esenciales para los estudios del cáncer y la respuesta a los medicamentos. Los análisis químicos y de proteínas se utilizan para evaluar el metabolismo de las células o tejidos, pero carecen de resolución unicelular e información espacial. Los ensayos basados en lectores de placas metabólicas pueden medir el pH y el consumo de oxígeno en la muestra a lo largo del tiempo y la posterior perturbación metabólica por productos químicos. El pH se puede utilizar para calcular la tasa de acidificación extracelular (ECAR), lo que proporciona una visión de la actividad glucolítica de las células9. Los métodos de imágenes de todo el cuerpo, incluida la tomografía por emisión de positrones fluorados-D-glucosa 2-[flúor-18] (FDG PET) y la espectroscopia de resonancia magnética (MRS), son modalidades de imágenes no invasivas utilizadas clínicamente para identificar la recurrencia tumoral y la eficacia del fármaco a través de mediciones metabólicas10,11,12,13,14.
FDG-PET toma imágenes de la captación tisular de FDG, un análogo de glucosa radiomarcado. El aumento de la captación de FDG-PET por los tumores en relación con el tejido circundante se debe al efecto Warburg12,13. MRS toma imágenes de núcleos comunes de moléculas utilizadas para el metabolismo, como 13C y 31P, y puede obtener información dinámica sobre cómo cambia el metabolismo en respuesta a estímulos, como el ejercicio o la alimentación14. Aunque FDG-PET y MRS se pueden usar clínicamente, estas tecnologías carecen de la resolución espacial para resolver la heterogeneidad intratumoral. Del mismo modo, las mediciones de consumo de oxígeno se realizan en una población masiva de células. Las imágenes de autofluorescencia superan el obstáculo de resolución espacial de estas tecnologías y proporcionan un método no invasivo para cuantificar el metabolismo celular.
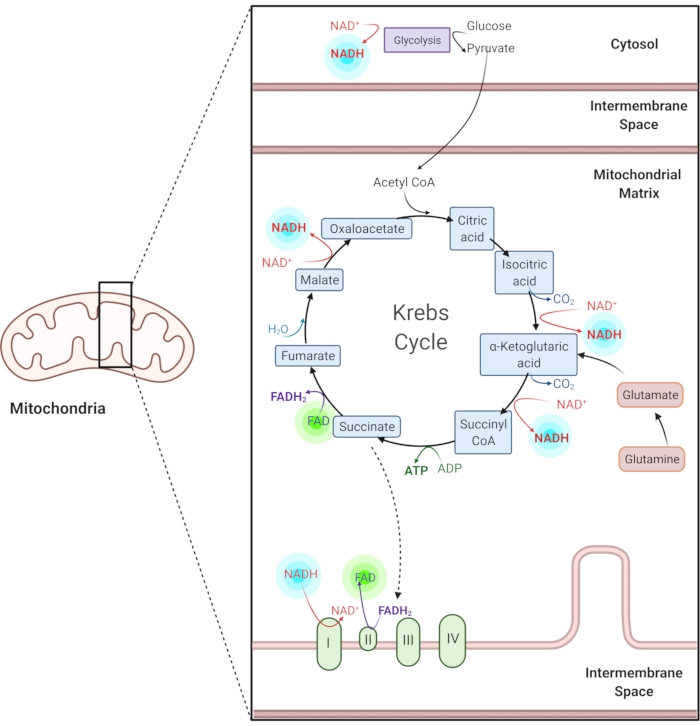
Figura 1: NADH y FAD en vías metabólicas comunes. NADH y FAD son coenzimas utilizadas en la glucólisis, el ciclo de Krebs y la cadena de transporte de electrones. Las imágenes de autofluorescencia de estas moléculas proporcionan información sobre el metabolismo celular. Haga clic aquí para ver una versión más grande de esta figura.
La nicotinamida reducida adenina (fosfato) dinucleótido (NAD(P)H) y el dinucleótido de flavina adenina oxidada (FAD) son coenzimas de reacciones metabólicas, incluyendo glucólisis, fosforilación oxidativa y glutaminólisis (Figura 1). Tanto EL NAD(P)H como el FAD son autofluorescentes y proporcionan contraste endógeno para la obtención de imágenes de fluorescencia1,15. NADPH tiene propiedades fluorescentes similares a NADH. Debido a esto, NAD(P)H se utiliza a menudo para representar la señal combinada de NADH y NADPH2,16.
Las imágenes de por vida de fluorescencia (FLIM) cuantifican la vida útil de la fluorescencia o el tiempo durante el cual un fluoróforo está en estado excitado. Las vidas útiles de fluorescencia responden al microambiente de los fluoróforos y proporcionan información sobre el metabolismo celular17. NAD(P)H y FAD pueden existir dentro de las células en conformaciones unidas a proteínas o libres, cada una de las cuales tiene una vida útil diferente. El NAD(P)H libre tiene una vida útil más corta que el NAD(P)H unido a proteínas; por el contrario, el FAD libre tiene una vida útil más larga que el FAD18,19 enlazado. Las vidas útiles y los pesos de los componentes de por vida se pueden cuantificar a partir de los datos de desintegración de la vida útil de la fluorescencia a través de Eq. (1)20:
I(t) = α 1e-t/τ1 + α 2e-t/τ2 + C (1)
Eq (1) representa la intensidad de fluorescencia normalizada en función del tiempo. Las α 1 y α 2 en esta ecuación representan los componentes proporcionales de vidas cortas y largas (α 1+ α 2=1), respectivamente, τ1 y τ2 representan las vidas cortas y largas, respectivamente, y C representa la luz de fondo7,20. La vida útil ponderada por amplitud, representada aquí como τm, se calcula utilizando Eq. (2).
τm= α 1τ1+ α 2τ2 (2)
Una vida media se puede calcular promediando “t” sobre la desintegración de intensidad del fluoróforo, que para una desintegración dos exponenciales se muestra por Eq. (3)17,21.
τ*m= (α 1τ12+ α 2τ22)/ (α 1τ1+ α 2τ2) (3)
Una imagen de intensidad de fluorescencia se puede calcular a partir de la imagen de por vida integrando la desintegración de la vida útil de fluorescencia. Las imágenes de autofluorescencia son un método no destructivo y libre de etiquetas que se puede utilizar para caracterizar el metabolismo de las células vivas a una resolución subcelular. La relación redox óptica proporciona una métrica analógica óptica del estado redox químico de la célula y se calcula como la relación de intensidades NAD(P)H y FAD. Aunque la fórmula para calcular la relación redox óptica no está estandarizada22,23,24,25, se define aquí como la intensidad de FAD sobre las intensidades combinadas de NAD(P)H y FAD. Esta definición se utiliza porque la intensidad sumada en el denominador normaliza la métrica entre 0 y 1, y el resultado esperado de la inhibición del cianuro es una disminución en la relación redox. Las vidas de fluorescencia de NAD(P)H libre y FAD proporcionan información sobre los cambios en el microambiente del disolvente metabólico, incluido el pH, la temperatura, la proximidad al oxígeno y la osmolaridad17.
Los cambios en la vida útil de fluorescencia de las fracciones unidas de NAD(P)H y FAD pueden indicar la utilización de la vía metabólica y el metabolismo específico del sustrato26. Los pesos de los componentes pueden interpretarse para los cambios en la fracción libre a la fracción unida de las coenzimas18,19. En conjunto, estas métricas cuantitativas de la vida útil de la autofluorescencia permiten el análisis del metabolismo celular, y la imagen de autofluorescencia se ha utilizado para identificar neoplasias de tejidos normales27,28, caracterizar células madre29,30, evaluar la función de las células inmunes31,32,33,34,35, medir la actividad neurológica36, 37,38, y comprender la eficacia de los medicamentos en tipos de cáncer como el cáncer de mama y el cáncer de cabeza y cuello21,39,40,41,42. Las imágenes de autofluorescencia de alta resolución se pueden combinar con la segmentación de imágenes para el análisis unicelular y la cuantificación de la heterogeneidad intrapoblacional43,44,45,46,47.
NAD(P)H y FAD se pueden obtener imágenes en microscopios de fluorescencia de fotón único o multifotón configurados para imágenes de intensidad o de por vida. Para los microscopios de fotón único, NAD(P)H y FAD se excitan típicamente en longitudes de onda de 375-405 nm y 488 nm, respectivamente, debido a fuentes láser comunes en estas longitudes de onda48. En la excitación de fluorescencia de dos fotones, NAD(P)H y FAD excitarán a longitudes de onda de aproximadamente 700 a 750 nm y 700 a 900 nm, respectivamente15,49. Una vez que los fluoróforos se excitan, NAD(P)H y FAD emiten fotones en longitudes de onda entre ~410 nm a ~490 nm y ~510 nm a ~640 nm, respectivamente15. Las longitudes de onda máximas de emisión NAD(P)H y FAD son de aproximadamente 450 nm y 535 nm, respectivamente48.
Debido a sus diferentes longitudes de onda de excitación y emisión, la fluorescencia de las dos coenzimas metabólicas puede aislarse espectralmente. Una comprensión de las características espectrales de NAD(P)H y FAD es necesaria para el diseño y la optimización de los protocolos de imágenes de autofluorescencia. El cianuro es un inhibidor del complejo IV de la cadena de transporte de electrones (ETC). Los efectos del cianuro sobre el metabolismo celular y las intensidades de autofluorescencia y la vida útil de NAD(P)H y FAD dentro de las células están bien caracterizados27,40. Por lo tanto, un experimento de perturbación de cianuro es un medio eficaz para validar los protocolos de imágenes NAD(P)H y FAD. Un experimento exitoso de cianuro proporciona confianza en que el protocolo de imágenes NAD(P)H y FAD se puede utilizar para evaluar el metabolismo de grupos o perturbaciones desconocidas.
