


ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
这些方案将帮助用户使用Seahorse细胞外通量分析来探测3D癌细胞系衍生球体中的线粒体能量代谢。
Abstract
近年来,三维(3D)细胞聚集体(称为球状体)已成为 体外 细胞培养的最前沿。与将细胞培养为二维单细胞单层(2D培养)相比,球状细胞培养促进,调节和支持 体内存在的生理细胞结构和特征,包括细胞外基质蛋白的表达,细胞信号传导,基因表达,蛋白质产生,分化和增殖。3D培养的重要性已在许多研究领域得到认可,包括肿瘤学,糖尿病,干细胞生物学和组织工程。在过去的十年中,已经开发了改进的方法来产生球状体并评估其代谢功能和命运。
细胞外通量(XF)分析仪已用于探索3D微组织中的线粒体功能,例如使用XF24胰岛捕获板或XFe96球状微孔板的球状体。然而,尚未详细描述使用XF技术探测球体中线粒体能量代谢的不同方案和优化。本文提供了使用球状微孔板和XFe96 XF分析仪探测单个3D球体中线粒体能量代谢的详细方案。使用不同的癌细胞系,XF技术被证明能够区分3D球体中的细胞呼吸,不仅具有不同的大小,而且具有不同的体积,细胞数量,DNA含量和类型。
使用寡霉素,BAM15,鱼藤酮和抗霉素A的最佳线粒体效应器化合物浓度来探测3D球体中线粒体能量代谢的特定参数。本文还讨论了对从球体获得的数据进行归一化的方法,并讨论了在使用XF技术探索球体代谢时应考虑的许多注意事项。该协议将有助于推动先进 体外 球体模型的研究。
Introduction
在过去的20年中,生物研究中体外模型的进步迅速发展。这些模型现在包括器官芯片模式,类器官和3D微组织球体,所有这些都已成为改善体外和体内研究之间转换的共同焦点。先进体外模型的使用,特别是球体,跨越多个研究领域,包括组织工程、干细胞研究、癌症和疾病生物学1,2,3,4,5,6,7,以及安全性测试,包括遗传毒理学8,9,10,纳米材料毒理学11,12、13、14、药品安全有效性检测8、15、16、17、18、19。
正常的细胞形态对生物表型和活性至关重要。将细胞培养成3D微组织球体允许细胞采用形态,表型功能和结构,更类似于 在体内 观察到的,但很难用经典的单层细胞培养技术捕获。 无论是在体内 还是在 体外, 细胞功能都直接受到细胞微环境的影响,其不仅限于细胞通信和编程(例如,细胞 - 细胞连接形成,形成细胞生态位的机会);细胞暴露于直接环境中的激素和生长因子(例如,细胞细胞因子暴露作为炎症反应的一部分);物理和化学基质的组成(例如,细胞是在坚硬的组织培养塑料中生长还是在弹性组织环境中生长);最重要的是,细胞代谢如何受到营养和氧气获取以及代谢废物(如乳酸)的加工的影响。
代谢通量分析是检查定义的 体外 系统中细胞代谢的一种有效方法。具体而言,XF技术允许分析完整细胞和组织的细胞生物能量学的实时变化。鉴于许多细胞内代谢事件发生在几秒钟到几分钟的量程内,实时功能方法对于了解 体外完整细胞和组织中细胞代谢通量的实时变化至关重要。
本文提供了使用强制聚集方法将癌症衍生细胞系A549(肺腺癌),HepG2 / C3A(肝细胞癌),MCF-7(乳腺癌)和SK-OV-3(卵巢腺癌)培养为 体外 3D球体模型的方案(图1)。它还 (i) 详细描述了如何使用 Agilent XFe96 XF 分析仪探测单个 3D 球体的线粒体能量代谢,(ii) 重点介绍了使用单个 3D 球体优化 XF 测定的方法,以及 (iii) 讨论了使用此方法探测 3D 球体代谢的重要考虑因素和局限性。最重要的是,本文描述了如何收集数据集,允许计算耗氧率(OCR)以确定氧化磷酸化,从而确定细胞球体中的线粒体功能。虽然没有针对该方案进行分析,但细胞外酸化率(ECAR)是XF实验中与OCR数据一起测量的另一个参数。但是,ECAR通常从XF数据集中解释得很差或不正确。我们针对按照技术制造商提供的基本方法计算 ECAR 的局限性进行了评论。
Subscription Required. Please recommend JoVE to your librarian.
Protocol

图 1:用于生成细胞球体、细胞外通量分析和下游测定的图形工作流程。 将四个癌细胞系选择性地培养为单层(A),从组织培养瓶中分离,并接种到超低附着物96孔微孔板中以形成球状体(B)。将A549肺癌、HepG2/C3A肝癌、SK-OV-3卵巢腺癌和MCF-7乳腺癌细胞接种于1×103-8×103 个细胞/孔,长大至7 d,形成单个球体,并通过连续观察和平面测量优化球体接种密度和培养时间。形成后,将单个球体洗涤到无血清XF培养基中,并小心地接种到球体测定微孔板中,并用聚-D-赖氨酸(C)预涂。使用XFe96分析仪对球体进行细胞外通量分析,使用几种方案来解决:(1)基础线粒体呼吸反应的最佳球体尺寸;(2)优化线粒体呼吸抑制剂滴定;(3)优化微孔板孔内球体放置。(D)使用XF后分析,相差显微镜和球体DNA定量进行数据归一化和其他下游 体外 测定。 请点击此处查看此图的大图。
1. 将癌细胞系培养为3D 体外 球状体
| 细胞系 | 描述 | 培养基 | 源 |
| 空客A549 | 肺癌细胞系 | 转速 1640 | 欧洲认证细胞培养物收集 (ECACC) |
| 丙酮酸钠(1 mM) | |||
| 青霉素- 链霉素 - (100 U/mL – 100 毫克/毫升) | |||
| 10 % (v/v) 断续器 | |||
| 七氢呋喃 2/丙三酸 | 肝癌细胞系,母体HepG2细胞系的克隆衍生物 | 断续器 | 美国组织培养收集 |
| 青霉素- 链霉素 - (100 U/mL – 100 毫克/毫升) | |||
| 10 % (v/v) 断续器 | |||
| 断续器7 | 乳腺癌细胞系 | 转速 1640 | 欧洲认证细胞培养物收集 (ECACC) |
| 丙酮酸钠(1 mM) | |||
| 青霉素- 链霉素 - (100 U/mL – 100 毫克/毫升) | |||
| 10 % (v/v) 断续器 | |||
| 断续器-3 | 卵巢腺癌细胞系 | 转速 1640 | 欧洲认证细胞培养物收集 (ECACC) |
| 丙酮酸钠(1 mM) | |||
| 青霉素- 链霉素 - (100 U/mL – 100 毫克/毫升) | |||
| 10 % (v/v) 断续器 | |||
| 元件 | RPMI 检测培养基(50 mL 终体积) | ||
| 基础介质 | 安捷伦海马 XF RPMI, pH 7.4 | ||
| 葡萄糖(1M无菌储备) | 11 mM(0.55 mL 储备溶液) | ||
| L-谷氨酰胺(200 mM无菌储备) | 2 毫升(0.5 毫升储备溶液) | ||
| 丙酮酸钠(100 mM无菌储备) | 1 毫升(0.5 毫升储备溶液) |
表1:癌细胞系培养基和XF培养基组成。
- 使用标准无菌组织培养技术培养所有细胞系,并使用合适的检测试剂盒确认它们不含支原体。
- 使用推荐的培养基在T75组织培养瓶或等效物中培养细胞系(表1)。将细胞系培养至65-80%汇合度,并定期传代至最多25次传代。
- 在杜贝克的改性磷酸盐缓冲盐水(DBPS)中冲洗细胞培养瓶两次。
- 用3mL细胞解离试剂(参见 材料表)在37°C下将细胞从烧瓶中分离5分钟,并通过显微镜确认分离。
- 轻轻吸出分离的细胞悬浮液以确保单细胞悬浮液,并用7mL完整组织培养基使细胞解离试剂失活。
- 通过以300× g 离心5分钟收集细胞,弃去上清液,并将细胞重悬于完全培养基中。
- 使用血细胞计数器或自动细胞计数器计数细胞,并滴定为接种所需的细胞密度。
注意:要以100μL/孔以4×103 个细胞/孔接种整个96孔板,应将细胞滴定至4×104 个细胞/ mL,推荐体积为12 mL。 - 将细胞悬浮液倾倒到无菌储液槽中,并使用多通道移液器将100μL细胞悬浮液分配到细胞排斥微孔的每个孔中。
注意:只有微孔板的内部60孔应接种,其余孔用DPBS填充。这将形成蒸发屏障,确保整个板的球体均匀性,并最大限度地减少板边缘效应。 - 将球状微孔板以300× g 离心15分钟,以迫使细胞进入松散的聚集体。
- 将板在37°C,5%CO2 下孵育至少3天以确保球状物形成。
- 使用标准化的实验室实践进行相差显微镜检查,以监测球体的生长。通过进行半体积培养基更换,每3天或每周补充两次细胞培养基。
2. 使用细胞外通量(XF)技术探测单个球体的线粒体能量代谢
- 检测准备(前一天)
- 使用相差为4倍放大倍率的倒置光学显微镜检查球体活力,以确保样品之间的完整球体结构,形态和整体均匀性。
- 为传感器墨盒补水。
- 将~20 mL口径滴入锥形管中。
- 将含有口径的锥形管置于非CO2 37°C培养箱中过夜。
- 从测定试剂盒中取出内容物。
- 从实用板上取下传感器滤芯,并将其倒置放在实用板旁边的工作台面上。
- 使用多通道P300移液管将200μL无菌ddH2O移液到传感器盒实用程序板的每个孔中。
- 将传感器盒放在公用设施板的顶部。
- 检查每个孔中的水位是否足够高,可以淹没传感器探头。
- 将组装好的传感器盒转移到非 CO2 37 °C 培养箱中,并保持过夜。
注意:此步骤可以在测定开始前12-72小时进行。
- 涂层球状体测定微孔板
- 使用无菌技术,将30μL /孔的无菌聚-D-赖氨酸(0.1mg / mL)溶液加入球状微孔板中,并在室温下孵育30分钟。
- 从球状微孔板的每个孔中吸出溶液,倒置板,然后将其牢固地敲击到薄纸上以除去任何残留的溶液。
- 用200μL /孔的无菌ddH2O洗涤板两次。
- 最后一次洗涤后,倒置微孔板并将其牢固地拍打到薄纸上以除去任何残留的水。
- 在使用前让板风干30分钟,或将其储存在4°C以备将来使用。
注意:球状体测定微孔板应涂有分子粘合剂,以确保球状物固定在微孔板的底部。如果没有分子粘合剂,球状体会脱落并干扰测定结果。其他分子粘合剂也可用作聚-D-赖氨酸的替代品,用于预涂板。预涂板可以储存在4°C,但在测定开始之前应保持平衡至室温。
- 制备XF测定培养基
- 制备XF RPMI培养基,如 表1所示,并使用0.22μm注射器过滤器进行无菌过滤器
- 测定准备(测定前1小时)
- 将补充的XF RPMI测定培养基预热至37°C。
- 将包被的球体测定微孔板预热在非CO2 37°C培养箱或干浴中。
- 准备传感器盒。
- 从空气培养箱中取出装有口径和传感器盒的锥形管。
- 从实用板上取下传感器盒,并将其倒置在工作台面上。
- 使用P300多通道移液器,从实用板中吸出水并将其丢弃。
- 将校准剂溶液倒入无菌试剂储液槽中,并使用P300多通道移液器将200μL/孔的预热校准剂加入实用板中。
- 拿起传感器滤芯并将其放回实用板的顶部,确保传感器完全浸没在校准液中。
- 将组装好的传感器滤芯移回非 CO2 37 °C 培养箱中,直到准备好装载端口注射液。
- 用测定培养基洗涤球状体。
- 从37°C,5%CO2 培养箱中取出球体培养板,并在显微镜下观察球体,以确保其在球体转移步骤之前的完整性。
- 用180μL/孔的预热测定培养基(包括任何背景校正孔)加载球状板的所有孔。
- 用3mL测定培养基部分填充7厘米培养皿。
- 使用装有宽孔口移液器吸头的多通道移液器,通过将移液器的抽吸体积设置为10-50μL,将球状体从96孔培养板转移到7cm培养皿中。
- 将种子球状体放入预包被的球状体测定微孔板中。
- 使用解剖显微镜和灯箱装置,将球体从培养皿转移到球体测定微孔板,如下所述。
- 将装有宽孔移液器吸头的单通道移液器的体积设置为20μL,并小心地吸出单个球体。将吸头直接置于球体测定微孔板的每个孔的中心,并允许重力将单个球体洗脱到每个孔的中心,即不要从移液器吸头中排出任何培养基,并允许毛细管作用从移液器吸头中取出球体。为了确认洗脱,可以在显微镜下将移液器的内容物移回7厘米培养皿中。
注意:单个球体的重力洗脱通常需要15-30秒,具体取决于球体的大小/密度。在此期间,不应取出移液器。任何背景校正孔都应不含球状体,并且仅含有测定培养基。在显微镜下,确认每个球体的位置。理想情况下,每个球体应位于每个孔的中心。 - 一旦所有球状体都转移到球状体测定微孔板中,在测定前将板转移到37°C的非CO2 培养箱中至少1小时。
- 将装有宽孔移液器吸头的单通道移液器的体积设置为20μL,并小心地吸出单个球体。将吸头直接置于球体测定微孔板的每个孔的中心,并允许重力将单个球体洗脱到每个孔的中心,即不要从移液器吸头中排出任何培养基,并允许毛细管作用从移液器吸头中取出球体。为了确认洗脱,可以在显微镜下将移液器的内容物移回7厘米培养皿中。
- 使用解剖显微镜和灯箱装置,将球体从培养皿转移到球体测定微孔板,如下所述。
3. 制备化合物并将其装载到XF测定的传感器盒中
| 注射策略 | 复合(端口) | XFe96 微孔起始体积 (μL) | 所需的终井浓度 | 端口体积(μL) | 最终 XFe96 微孔体积注射后 (μL) | 工作库存集中 |
| 1 | 寡霉素 (A) | 180 | 3微克/毫升 | 20 | 200 | 30微克/毫升 |
| 鱼藤酮 (B) | 200 | 2 微米 | 20 | 220 | 22 微米 | |
| 抗霉素 A (B) | 200 | 2 微米 | 20 | 220 | 22 微米 | |
| 2 | 巴姆15 (A) | 180 | 5 微米 | 20 | 200 | 50 微米 |
| 鱼藤酮 (B) | 200 | 2 微米 | 20 | 220 | 22 微米 | |
| 抗霉素 A (B) | 200 | 2 微米 | 20 | 220 | 22 微米 |
表2:使用XFe96分析仪探测单个3D球体的线粒体能量代谢的线粒体化合物浓度。
- 使用完全补充的预热XF RPMI测定培养基制备 表2 中所述的每种化合物的工作储备浓度。
- 按列方向定位墨盒板(与实用板耦合),从左到右 1-12。
- 如果使用装载导轨,请根据装井程序将其放置在墨盒板上,例如,如果首先装载端口A,请确保 A 在导轨的左上角可见。
- 将每种化合物的工作溶液转移到合适的储液槽中,并使用校准的P100多通道移液器将20μL分配到所有相应的端口中。对每个化合物重复上述步骤,将其放入其余端口中。
注意:如果传感器盒板上未使用任何端口,则可以将其留空或填充测定培养基。如果只选择特定的端口号,请确保与该字母对应的其他端口装载了测定介质;否则,空气将被注入井中,从而影响这些井中的结果。 - 端口加载后,取下板加载导轨(如果使用)并准备分析仪以加载传感器盒。
注意:如果在加载端口后未立即进行测定,请将盖子放回传感器盒上,并将板放回37°C空气培养箱中,直到准备好装入机器。
4. 分析设计、注射策略和数据采集
- 运行检测
- 打开分析仪电源并连接到控制器(计算机)。
注:这可以通过波形控制器软件小部件面板中的仪器连接状态进行验证。 - 导航到WAVE软件中的 模板 页面,找到实验的检测模板文件,然后双击将其打开。
注意:如果检测模板未出现在 “模板” 视图中,请将模板文件从共享网络驱动器或 USB 闪存驱动器导入模板文件夹。 - 要开始检测,请单击 运行检测 选项卡。
注意:如果在板图中正确分配了组定义,则分析将准备好按照页面右侧绿色勾号所示运行。在此阶段,可以在测定摘要页面或空白页面上输入任何其他信息;继续执行下一步。由于线粒体调制器在3D微组织球体中的延迟穿透(图2),请使用 表3中描述的测量方案信息。
- 打开分析仪电源并连接到控制器(计算机)。
| 测量周期 | 注入编号和端口 | 测量细节 | 周期持续时间(小时:分钟:秒) |
| 校准 | 不適用 | XF分析仪始终执行此校准,以确保测量准确 | 00:20:00(这是平均值,可能因机器而异) |
| 平衡 | 不適用 | 校准后发生平衡,建议使用。 | 00:10:00 |
| 基底 | 不適用 | 周期 = 5 | 00:30:00 |
| 混合 = 3:00 | |||
| 等待 = 0:00 | |||
| 测量 = 3:00 | |||
| 寡霉素/BAM15 | 注入 1(端口 A) | 周期 = 10 | 01:00:00 |
| 混合 = 3:00 | |||
| 等待 = 0:00 | |||
| 测量 = 3:00 | |||
| 鱼藤酮 + 抗霉素 A | 注入 2(端口 B) | 周期 = 10 | 01:00:00 |
| 混合 = 3:00 | |||
| 等待 = 0:00 | |||
| 测量 = 3:00 | |||
| 总时间: | 03:00:00 | ||
表 3:使用 XFe96 分析仪探测单个 3D 球体的线粒体能量代谢的实验方案设置。
- 单击 “开始运行 ”以显示 保存位置 对话框。
- 输入结果文件的 保存位置 ,然后将组装好的传感器盒放在分析仪侧面门上显示的热托盘上。等待热托盘自动打开,并等待屏幕显示 “装载校准器”实用程序板 消息。在按照屏幕上的提示操作之前,请确保 i) 传感器墨盒正确安装在实用板上,ii) 盖子已从传感器盒上取下,以及 iii) 传感器盒在实用板上的正确方向。
- 按照屏幕上的命令启动传感器盒校准。
注意:完成校准所需的时间约为10-20分钟(用于37°C下的测定)。 - 传感器盒校准后,按照波形控制器上的屏幕说明将球形微孔板加载到分析仪中,以启动12分钟的平衡步骤。
注意:带有白色勾号的绿色框表示该孔的“良好”校准。如果任何孔未能提供“良好”校准,则将用红色框和白色十字表示。在使用修饰测定选项卡完成测定后,应注意此类孔并将其排除在任何 分析 之外。 - 等待分析仪在机器完成平衡步骤后自动开始采集基线测量值(如仪器协议中所述)。
- 要完成实验,请按照 WAVE 控制器上的屏幕命令进行操作。
注意:从分析仪中取出球状微孔板后,丢弃传感器盒,并留出球状板,以便在必要时进行进一步分析(例如,双链(ds)DNA定量)。如果进一步分析不需要微孔板,可以将其与传感器盒一起丢弃。 - 等待分析对话框出现并查看结果或返回 模板 视图。
5. 数据归一化和分析策略 - 分析后归一化和下游分析(可选步骤)
- 数据规范化
- 要对球体数据进行归一化,请参阅与数据归一化策略相关的一系列方案,用于计算球体大小和体积以及量化球体测定中的 dsDNA。这些已作为补充文件列入;请参阅 补充文件 1 和 补充文件 2。
- 数据分析
- 要将数据导出到其中一个自动分析发生器中,请按照WAVE控制器上的数据导出命令进行操作,然后选择与测定类型匹配的导出发生器。或者,导出数据文件并将其上传到海马分析中。
注意:报告生成器和Seahorse分析的缺点是数据分析仅限于XF分析的设计方式,并且不允许在整个测量周期中取平均值。从仪器软件手动导出数据集允许用户在这方面偏好。鉴于用于评估3D球体的线粒体呼吸的注射策略可能与典型的“MitoStress”测试不同,因此已经开发了一系列电子表格模板来帮助分析这些数据集,特定于3D细胞培养物,并将根据要求提供。这些数据模板文件将提供有关关键线粒体呼吸参数的数据,详见 图2并加以说明。 - 要分析数据,请从WAVE控制器软件将数据导出为电子表格报告,并使用独立的电子表格模板进行分析。
- 要将数据导出到其中一个自动分析发生器中,请按照WAVE控制器上的数据导出命令进行操作,然后选择与测定类型匹配的导出发生器。或者,导出数据文件并将其上传到海马分析中。

图 2:从细胞外通量数据分析派生参数的示意图描述符。 简称:OCR = 耗氧率。 请点击此处查看此图的大图。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
为了获得形状良好的致密球体,每个细胞系分别针对接种密度和培养持续时间进行了优化(图3)。A549,HepG2 / C3A和SK-OV-3细胞系最初形成松散的聚集体,直到培养7天后才进展为具有明确定义周长的圆形球状体。相反,MCF-7细胞可以在3天内形成球状体。所有球体模型的初始细胞接种密度与培养期后的球体体积之间存在明显的相关性。根据接种密度优化了球体尺寸和形态。在所有模型中,随着球体尺寸的增加,形态和圆度开始下降。在A549和SK-OV-3细胞的4×103 个细胞/孔中优化细胞系的接种策略;HepG2 / C3A细胞先前已在其他地方优化为1×103 个细胞/孔,并且在所有测定中以4×103 个细胞/孔使用MCF-7细胞。在优化的接种策略下,球体体积在5.46×107μm3 (SK-OV-3)和1.45×108μm 3 (A549)之间(图3B)。所有球体类型的初始晶种密度与球体体积之间均呈线性相关,其中A549和HepG2/C3A的R2 值分别为0.957和0.947。MCF-7和SK-OV-3球体体积均与初始接种密度R2 = 0.977具有更大的相关性(图3A)。
使用FIGUI分析软件中的图像平面测量法使用长球体直径和短球体直径计算球体圆度。完美的旋转椭球对称性具有圆度= 1.0;偏离1.0表示圆度损失(图3C)。MCF-7球体的圆度大于其他模型,在所有晶种密度下,圆度保持在0.83和0.9之间。相比之下,SK-OV-3球体的外周没有明确界定,即使在培养7天后,球体体积也明显变小,在4×103/孔的接种密度下,产生最大圆度为0.61的球体。还发现HepG2 / C3A细胞形成紧密,形成良好的球状体,在所有球体的表面积上具有均匀的形态,对于以1×103 个细胞/孔接种的细胞,圆度保持在0.79。A549细胞似乎遵循一种趋势,其中球状圆度和形态随着接种密度的增加而增强;然而,在这些实验中使用的密度下,圆度不大于0.63。
基础线粒体呼吸计算为OCR从接种在超低附着球体培养微孔板中的1×103,2×103,4×103或8×103个细胞/孔的球体中测量(图3D)。对于所有球体类型,OCR随球体大小增加而增加,并且与球体体积线性相关,其中MCF-7球体中的R2最高,为0.988,SK-OV-3球体中最低,为0.744(图3E)。所有实验组之间的测量OCR在统计学上都不同。A549具有最低的OCR,在最大的球体尺寸下仅达到18 pmol /分钟/孔(图3D)。相反,MCF-7球体在培养仅3天后,在最小的球体尺寸下产生类似的OCR,对于最大的球体尺寸,达到53 pmol /min /孔的最大基线OCR(图3D)。HepG2/ C3A产生的OCR数据与球体大小和形态高度一致。在从1个×10个3个细胞/孔接种的HepG2 /C3A球体中,基线OCR平均达到15 pmol /分钟/孔,在最大的球体中增加到最大52 pmol /min /孔(图3D)。SK-OV-3球状体的OCR仅在从4个×10 3个细胞/孔和8个×103个细胞/孔生长的球体之间显着,在1×103,2×103或4×103个细胞之间生长的球体差异不大。尽管大小不同,但所有尺寸点的HepG2 / C3A和MCF-7球体之间的OCR数据都高度相似。相对于球体大小(μm3),MCF-7球体的基线OCR与每孔1,000个细胞在7天内生长的HepG2 / C3A球体相当。

(A)A549,HepG2 / C3A,MCF-7和SK-OV-3球体针对接种密度进行了优化,并分别在1×103,2×10 3,4×103和8×103细胞/孔下监测其生长;比例尺 = 500 μm。(B)使用收集的显微照片的平面数据计算球体尺寸,并使用Pearson的相关统计量进行比较;虚线表示 95% 置信区间的分布。(C)通过计算圆度来比较球体形态。(D)测量5x OCR,之后使用安捷伦海马XFe96分析仪添加鱼藤酮 - 抗霉素A以考虑非线粒体呼吸频率。将 OCR基础测量为 OCR - OCRr/a 在接种密度 (D) 和球体体积 (E) 之间进行比较。数据是每个球体类型和细胞接种密度从5-8孔重复±SEM的平均值。简称:OCR =耗氧率;OCR基础=基础线粒体呼吸;OCRr/a = 添加鱼藤酮-抗霉素A后的OCR.请点击此处查看此图的大图。
在XF分析中暴露于呼吸调节剂的浓度和时间过程是测定优化的关键步骤。将呼吸调节剂化合物 - 寡霉素,BAM15,鱼藤酮 - 抗霉素A-或DMSO载体对照的混合物依次通过传感器盒注射口注射到含有MCF-7球体的微孔板孔中(图4A)。完成四个测量周期,以确定所有样品组的平均基础OCR为30-40 pmol /min /孔。在测定持续时间的剩余时间内,每5个测量周期依次加入呼吸调节剂,以达到注射1后0.5μM(0.5μg/ mL寡霉素)的最终孔浓度;注射后2.0微米(2微克/毫升寡霉素)2;注射后5微米(5微克/毫升寡霉素) 3;最后,第四次序贯加药后最大孔浓度为11μM(11μg/mL寡霉素)。在整个实验过程中,MCF-7球体对车辆控制没有反应(图4A)。基础OCR在第一次以0.5μM或0.5μg/mL寡霉素的最低浓度注射每种化合物后立即发生变化(图4B)。在第一次注射0.5μg/mL后5个测量周期后,用寡霉素将MCF-7球状体的OCR从41 pmol/min/孔降低到23 pmol/min/孔(图4B)。
响应于0.5μm BAM15,在第二次进样前将OCR从33升高至41 pmol /分钟/孔(图4C)。相比之下,鱼藤酮加抗霉素A的组合在第二次注射前将OCR从37 pmol / min /well降低到13 pmol / well(图4D)。动力学痕迹进一步显示OCR中稳定的线性减少(寡霉素和鱼藤酮 - 抗霉素A)或增加(BAM15)。对于所有化合物给药方案,在10-12个完整的测量周期(60-72分钟)内,在总孔浓度为2μM BAM15,2μM鱼藤酮,2μM抗霉素A和2μg/ mL寡霉素的情况下实现稳态OCR(图4A)。耗氧速率在~19匹马/分钟/孔(寡霉素),52匹马/分钟/孔(BAM15)和10匹马力/分钟/孔(鱼藤酮-抗霉素A)(图4A)时达到稳态平台。进一步增加寡霉素,BAM15或鱼藤酮加抗霉素A的化合物浓度对OCR没有明显影响,OCR在测定的其余部分保持不变。这些数据表明,使用3D球体时,应考虑化合物浓度和暴露于呼吸调节剂化合物的时间过程以进行测定优化。

图4:将呼吸调节剂化合物滴定作为优化细胞外通量分析的重要步骤 .(A)将MCF-7球体接种在4×10个细胞 /孔中并培养3天,然后放入含有XF RPMI的球形测定微孔板的孔中,并使用XFe96分析仪探测OCR±线粒体调节剂。测量5x OCR,之后滴定载体对照,少霉素(B),BAM15(C)或鱼藤酮 - 抗霉素A(D)以分别抑制线粒体ATP合酶,确定最大呼吸能力或建立非线粒体呼吸频率。在4个单独的滴定注射策略(0.5μM,1.5μM,3μM和6μM,寡霉素单位为μg/ mL)中增加每个线粒体调节剂的浓度,以确定响应最佳化合物浓度的最大稳态OCR。在每次注射之间测量OCR5个测量周期。数据是来自5-8个井重复±SEM的平均值。简称:OCR = 耗氧率。 请点击此处查看此图的大图。
XF技术的主要优点之一是能够探测完整细胞和组织中的线粒体功能。为了检查细胞和组织中线粒体功能的特定方面,线粒体调节剂通过传感器盒上的4个可用进样口依次添加到样品微孔板的孔中。在XF测定中用于探测线粒体参数的典型调节剂序列是寡霉素,质子团(例如FCCP或BAM15)和鱼藤酮加抗霉素A的组合,它们依次添加以抑制线粒体ATP合酶,确定最大呼吸能力,并分别校正非线粒体呼吸频率。这种典型的调制器添加序列被测定技术制造商称为线肌压力测试。鉴于寡霉素可以抑制某些细胞单层20中的解耦刺激呼吸,我们通过在(单次)和(顺序)寡霉素注射之前(单次)和之后测量非耦合刺激的OCR(OCRmax)来检查癌症衍生的3D球体(图5A-D)。OCRmax没有显着限制在由HEPG2 / C3A或SK-OV-3形成的球体中添加寡霉素(图5E和图5G)。然而,与单次注射BAM15获得的OCR最大值相比,在寡霉素后连续注射BAM15后,A549和MCF-7球体的OCRmax显着降低(图5F和图5H)。除非另有说明,因此建议使用单独的孔用寡霉素和解偶联剂治疗,在探索3D球体的线粒体能量代谢时最终加入鱼藤酮和抗霉素A。这种方法仍然允许计算所有线粒体参数,就像典型的线肌压力测试一样,其中化合物是按顺序添加的。

图5:线粒体呼吸化合物的单次或连续注射。将 MCF-7、HEPG2/C3A、SK-OV-3 和 A549 的癌细胞衍生球体放入 XF RPMI 中 XFe96 球状微孔板的孔中,并使用安捷伦海马 XFe96 分析仪探测 OCR。测量5x OCR,之后分别测定2 μg/mL寡霉素(注射口A:绿色痕量)或5 μM BAM15(注射口A:蓝色迹线或注射口B:绿色迹线)以抑制线粒体ATP合酶并确定最大呼吸能力。动力学 OCR 数据以基础百分比 (A-D) 表示。最大呼吸能力(OCR最大值)通过以下公式计算为基础OCR的因子:OCR最大值= OCRBAM15 / OCR基础。 OCRmax是从BAM15注射后8-10个测量周期的OCR平均值获得的,有(绿色条)和没有(蓝色条)寡霉素。数据是来自球体测定微孔板中3-8个个体孔重复的SEM±平均值。缩写:OCR = 耗氧率。请点击此处查看此图的大图。
利用这些优化实验中确定的最佳细胞接种密度、化合物浓度、注射策略和测量周期(表3),我们制定了精确探测基础线粒体呼吸的详细方案:OCR基础 呼吸(图6A),ADP磷酸化呼吸:OCRADP (图6B),泄漏呼吸:OCRomy (图6C),耦合效率(图6D)),最大呼吸能力:最大 OCR(图6E)和备用呼吸能力:使用癌症衍生的3D球体进行OCR备用 (图6F)。

图6:使用XF技术探测OCR以建立癌症衍生球体的线粒体能量代谢。将 MCF-7、HEPG2/C3A、SK-OV-3 和 A549 的癌细胞衍生球体置于 XF RPMI 中球体测定微孔板的孔中,并使用安捷伦海马 XFe96 分析仪探测 OCR。测量5x OCR,之后加入2μg/mL寡霉素或5μM BAM15,并加入RA以抑制线粒体ATP合酶,确定最大呼吸能力,并分别计算非线粒体呼吸频率。(A)基础线粒体呼吸(OCR基础)计算为端口A注射前3个测量周期的OCR平均值。(B)氧化磷酸化的偶联效率通过表示OCRADP(OCR基础 - OCR泄漏)作为OCR基础的百分比来近似。(C)将ADP磷酸化呼吸(OCRADP)测量为寡霉素敏感的OCR,根据BAM15注射前11-13个测量周期的平均OCR计算得出。(D)将渗漏呼吸(OCR泄漏)测量为对寡霉素不敏感的OCR,根据测量周期11-13的平均OCR计算。(E)最大呼吸能力(OCR最大值)被测量为BAM15注射后测量的平均最大OCR。(F) 通过表示 OCR备用(OCR最大值 - OCR基础)作为 OCR 基础的百分比来计算备用呼吸能力。从所有速率中减去鱼藤酮 - 抗霉素A注射后的OCR以校正非线粒体OCR。数据是来自XFe96球状板3-8个单个孔重复的SEM±平均值。简称:OCR =耗氧率;RA = 2 μM 鱼藤酮-2 μM 抗霉素 A.请点击此处查看此图的大图。
使用在3天内从4×103 个细胞/孔中生长的MCF-7球状体作为模型,以确定球状体测定微孔板内的最佳转移,放置和分析。使用制造商为球状微孔板提供的尺寸,将孔表面分为三个区域,以实现最佳的球体放置(图7A),其中区域1被突出显示为孔中心的最佳区域。使用宽孔移液器吸头小心移液,将球状体转移到球状板中,并通过重力洗脱随机分布在孔面上(图7B)。在使用重力洗脱小心转移球体的情况下,大多数球状体通常可以使用制造商推荐的转移技术在微孔板的区域1-2中找到。在通过抽吸将球体从移液器吸头中挤出的地方,球状体通常放置在这些区域之外,并且无法使用显微镜看到。
为了比较球体放置位置,将MCF-7球体转移到指定区域1-3或区域外的球状体测定微孔板中(图7A)。在基线时和加入寡霉素,BAM15或鱼藤酮 - 抗霉素A之后,通过动力学实验OCR跟踪这4个孔(图7C)。OCR是根据每次注射前三个周期读数的平均值计算的(图7B)。在4个选定的孔中以200分钟以上的动力学测量OCR(图7C)并进行基线校正(图7D)。当旋转椭球体放置在区域3或区域外时,基线OCR明显低于放置在区域1和2中的旋转椭球体(图7C)。与区域3和区域外区域相比,位于区域1和2的球体之间,呼吸化合物寡霉素,BAM15和鱼藤酮 - 抗霉素A的作用也存在显着差异。在放置在区域3或区域外的球体中观察到寡霉素的OCR增加(图7E)。此外,放置在区域3或区域外的球体对BAM15的反应过高,并且在鱼藤酮 - 抗霉素A注射后OCR高于基线(图7E)。尽管将球体放置在区域2与区域1的基底OCR(图7C)几乎增加了两倍,但对所有呼吸化合物的响应的折叠变化非常相似(图7E),这表明放置在区域1或2中的球体之间的基础OCR差异不太可能是放置在孔内的结果。

图 7:在球体测定微孔板中放置球体决定了使用 XF 技术的基础 OCR 和线粒体调节剂效应。 将MCF-7球状体接种在4×103 个细胞/孔中,培养3天,然后放入含有XF RPMI的球状微孔板的孔中,并使用安捷伦海马XFe96分析仪探测OCR±线粒体调节剂。(A)测定持续时间后球体测定微孔板中球体区域位置的显微照片;比例尺 = 500 μm,并且随着时间的推移从相应的孔中捕获的 OCR 表示为 pmol/min-1/孔-1 (B) 或 % 基础 (C)。(D)将MCF-7球体置于球体测定微孔板内不同区域的线粒体调节剂效应;数据表示为基底的褶皱变化。(E)示例动力学迹线突出显示使用OCR数据测量(红色圆圈)来计算每个线粒体调制器对 E中呈现的数据的响应。显示的数据来自单个油井响应。简称:OCR = 耗氧率。 请点击此处查看此图的大图。
背景的选择标准非常重要;使用最外层的孔进行背景校正并不代表所有微孔板孔,这可能导致由于球体微孔板的边缘效应而得出不正确的数据假设和错误的数据结论。为了评估这一观察结果,使用MCF-7球体来比较测定校正程序,以得出响应于添加载体对照,寡霉素,BAM15或鱼藤酮 - 抗霉素A的OCR值(图8)。所有呼吸化合物都产生了所选化合物的预期动力学OCR谱,揭示了平均稳定的基础呼吸速率为20-30 pmol / min /孔(图8A)。然而,使用最外层的孔进行背景温度校正(A1,A12,H1和H12)分析测定数据时,添加呼吸化合物后OCR显示的值特别低;OCR对鱼藤酮-抗霉素A产生负值。为了响应这些观察结果,使用一系列随机分布在球体微孔板上的空孔作为背景温度校正孔进行替代分析(图8B)。在应用替代背景校正的情况下,两个分析集中对OCR的所有相对复合效应都是相同的;然而,绝对OCR值增加了约10 pmol /分钟/孔(图8)。这些数据强调了背景温度校正对球体测定微孔板的强大功能和重要性,并强调了用户优化XF分析的重要性。

图 8:随机选择用于背景校正的孔,以改善对球体测定微孔板温度梯度的控制。 OCR数据从 图2A 推断,使用推荐的背景校正孔(A)与随机分配的背景校正孔(B)。简称:OCR = 耗氧率。 请点击此处查看此图的大图。
与细胞单层不同,球体代表3D空间中细胞的异质聚集,因此需要在分析方面进行彻底的考虑,特别是在对这些数据进行归一化时。本文介绍了三种对从MCF-7旋转椭球体获取的XF数据进行归一化的方法(图9)。当非规范化时,当与Pearson相关系数(P = 0.0057)进行统计学比较时,OCR与球体大小(由初始细胞接种密度决定)显着正相关(R 2 = 0.98)(图9A)。当OCR归一化为初始细胞接种密度(R2 = 0.78)时,这种线性关系降低,并且不再与球体大小显着相关(P = 0.117,图9B)。当归一化为球体体积(R2 = 0.77;皮尔逊相关系数P = 0.120,图9C)和核dsDNA含量(R2 = 0.58;皮尔逊相关系数 P= 0.233,图 9D)。这些数据强调了在探测球体的线粒体代谢时对XF数据进行归一化的重要性,特别是如果它们的大小不同。

(A)从MCF-7培养3天中获得的原始OCR数据,并使用Pearson模型绘制,以获得球体接种密度与OCR之间的相关系数;P 值设置为 0.05。(B) 原始OCR数据根据初始球体播种密度进行归一化;(C)从显微镜平面测定法获得的MCF-7球体体积;和(D)核ds DNA含量比较使用皮尔逊相关系数。简称:OCR =耗氧率;ds 基因 = 双链基因。请点击此处查看此图的大图。
| 细胞系 | 播种密度(井) | 球状生长(天) | 最终球体体积 (μM3) | 基础口服补液(吡米洛2/分钟/孔) | 满足基础 OCR 的灵敏度(是/否) |
| 斯科夫 | 1000 | 5 | 9.52E+06 | 28 ± 3.5 | 是的 |
| 斯科夫 | 2000 | 5 | 2.38E+07 | 26 ± 1.4 | 是的 |
| 斯科夫 | 4000 | 5 | 4.92E+07 | 36 ± 3.1 | 是的 |
| 斯科夫 | 8000 | 5 | 1.11E+08 | 50± 7.9 | 是的 |
| 七氢呋喃G2 | 1000 | 5 | 1.11E+07 | 15 ± 0.7 | 不 |
| 七氢呋喃G2 | 2000 | 5 | 2.88E+07 | 23 ± 1.8 | 是的 |
| 七氢呋喃G2 | 4000 | 5 | 5.46E+07 | 31 ± 1.7 | 是的 |
| 七氢呋喃G2 | 8000 | 5 | 1.21E+08 | 52± 2.8 | 是的 |
| 空客A549 | 1000 | 5 | 2.11E+07 | 30 ± 2.5 | 是的 |
| 空客A549 | 2000 | 5 | 3.57E+07 | 41 ± 1.6 | 是的 |
| 空客A549 | 4000 | 5 | 6.93E+07 | 53± 7.2 | 是的 |
| 空客A549 | 8000 | 5 | 1.44E+08 | 65± 8.4 | 是的 |
| 断续器-7 | 1000 | 3 | 1.60E+07 | 29 ± 0.8 | 是的 |
| 断续器-7 | 2000 | 3 | 2.52E+07 | 37 ± 1.7 | 是的 |
| 断续器-7 | 4000 | 3 | 6.00E+07 | 46 ± 1.7 | 是的 |
| 断续器-7 | 8000 | 3 | 1.06E+08 | 66 ± 2.9 | 是的 |
表 4:用于确定单个 3D 椭球体中基础 OCR 测量值的优化参数。 简称:OCR = 耗氧率。
补充文件1:分析球体大小和体积。请按此下载此档案。
补充文件2:从球状微孔板中的球状体中定量双链DNA。请按此下载此档案。
补充文件3:关于获得可靠的XF测定数据集所需的重复次数的建议。请按此下载此档案。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
二. 主要结论和产出
本文提供了使用XFe96 XF分析仪的一系列癌症衍生细胞系探测单个3D球体的线粒体能量代谢的详细方案。开发并描述了一种利用细胞排斥技术强制聚集快速培养A549、HepG2/C3A、MCF7和SK-OV-3细胞球体的方法。该协议解决了使用XF技术探测球体代谢的许多考虑因素,包括(1)球体培养方案的优化以及将球体处理和转移到技术制造商从其原始培养容器中分离到特定球体测定微孔板中;(2)要使用的呼吸化合物的浓度和化合物渗透的时间依赖性;(3)注射策略要采用;(4)在实验组之间规范化数据的方法。所有这些考虑因素已在本文件中进行了审查,并将在下文中进一步详细讨论。这些方法被介绍为使用XFe96通量分析仪的单个3D球体生成一致代谢氧通量数据的简化方法。这种实验方法可以用作起点和量规,用于其他在基本实验室环境中轻松实现的球体模型。
考虑
XF技术的球体生长、大小和灵敏度
为了使用XF技术建立可重复的数据,必须针对特定模型表征和优化测定。这种方法在基本的细胞单层中相对简单;然而,这在将细胞培养为3D球体时带来了额外的挑战。在这里介绍的实验中,制造商的RPMI培养基在使用时进行了补充。虽然注意到一些细胞系,即HepG2 / C3A,在DMEM生长培养基中培养,但在这些相对较短的测定期间(约3-5小时),用RPMI-DMEM制剂取代对XF分析的影响有限。这两种培养基的配方非常相似,用户可以通过补充来“调整”Seahorse RPMI培养基以匹配其细胞培养基的基质,例如,增加葡萄糖,进一步添加碳水化合物来源。对于所有XF缓冲液和培养基的最终配方至关重要的是没有酚红,这可能会干扰XF探针盒板内的荧光探针和碳酸氢钠,碳酸氢钠会导致碱性,因为细胞培养箱中存在CO2 缓冲。其他介质和缓冲液可以在内部购买和/或制造。例如,克雷布斯·林格HEPES缓冲液是一种简单的缓冲液,可用于评估许多不同细胞的呼吸,包括球体模型。然而,XF测定的用户应注意,培养基/缓冲液及其补充的变化可能会改变其整体缓冲能力。当用户可能对测量ECAR感兴趣时,这尤其令人担忧,其中需要评估培养基的缓冲因子以允许ECAR转化为质子流出速率(PER)。
由于当孔中的细胞数量在系统的灵敏度范围内时,XF技术测量的细胞OCR与细胞密度成正比,因此使用单个3D球体研究这种关系非常重要。通过探测从每孔1,000,2,000,4,000或8,000个细胞密度接种的4个不同癌细胞系培养的单个3D球体的OCR,我们表明XFe96分析仪足够灵敏,可以拾取从不同细胞接种密度生长的3D球体之间的线粒体呼吸速率的变化(图3)。我们表明,细胞接种密度的最佳范围,以及因此形成用于探测OCR的3D球体的球体体积,因细胞类型而异。这可以通过OCR与晶种密度或球体体积之间的线性关系来表示(图3)。对于A549和HepG2 / C3A细胞,OCR灵敏度的最佳接种密度在1,000至8,000个细胞/孔之间;MCF-7为2,000-8,000个细胞/孔,SK-OV-3细胞为4,000-8,000个细胞/孔。这些数据表明,在使用XF技术评估OCR时,球体尺寸的优化尤为重要。
关于最小和最大球体体积以及基础 OCR 的注意事项
通常,制造商为这些实验推荐的可测量OCR参数始终存在最小和最大阈值。对于XFe96分析仪,20 pmol O2/分钟/孔和200 pmol O2/min/孔之间的基础OCR分别是下限和上限。单层细胞和球状体就是这种情况,实验模型位于该动态OCR范围内的位置将取决于可用的生物材料的数量,例如,作为单层的细胞数量或球体的大小。有关此处使用的旋转椭球体模型如何实现 OCR 阈值的示例,请参见 表 4 。谨慎的做法可能是检查油井内的氧含量,这些数据也可以从这些测量值中作为液位数据获得。出于质量控制目的,应从每个实验中常规查看。如果井中存在氧气消耗,这将在数据中显现出来。如果是这种情况,可能需要调整实验中的测量周期;例如,增加混合步骤使得在测量周期内的下一个测量周期之前恢复井中的氧含量。虽然有可能,但我们发现这对于使用所述细胞系的单球体实验来说非常不可能。
选择线粒体解耦合剂进行细胞外通量测定
质子离子载体,如羰基氰化物4-(三氟甲氧基)苯腙(FCCP)21,羰基氰化物间氯苯基腙(CCCP)22 或BAM1523,是有效的小分子化学物质,能够破坏线粒体膜上的电化学质子梯度,抑制ATP的产生,并最终解开线粒体呼吸24.新的小分子继续为这些目的而开发,特别是在治疗代谢疾病25,26,27;参考两个优秀的评论28,29。相反,氧化呼吸的解偶联与不希望的脱靶毒性30有关。然而, 在体外 细胞测定中,分子FCCP去极化线粒体膜电位并发挥脱靶效应,例如质膜去极化,破坏NA + 离子通量31;干扰细胞蛋白处理32,甚至诱导细胞衰老33。BAM15最初于2013年作为线粒体解耦合器引入,对质膜23的影响最小,质子活性在整个细胞的微摩尔范围内,在分离的线粒体23,34中具有纳米摩尔范围。
鉴于FCCP对质膜去极化的效力,BAM15是一种更可靠的质子团,用于在细胞外通量测定中解偶联完整全细胞中的呼吸。尽管FCCP及其对应物CCCP已被用于测定最大呼吸能力超过50年,并继续广泛用于XF研究,但这些小分子的使用往往低估了线粒体和细胞代谢能力。这在一定程度上与为什么这么多使用XF技术的出版物落入报告负备用呼吸能力的陷阱或低估使用FCCP时的真实线粒体呼吸能力有关。FCCP在完整细胞和组织中添加的效力通常会导致线粒体功能受损,并且细胞可能难以在添加后的多个测量周期内适当地操作以维持最大呼吸能力,即使在非常低的浓度下也是如此35。因此,在许多研究中可以发现细胞对FCCP的反应在初始测量周期周期后下降。虽然FCCP通常用于XF分析,但BAM15优先用于涉及整个细胞或球体模型的情况,因为它可以在浓度高达10μM 3的完全去极化线粒体中保持最大的呼吸能力。此外,BAM15诱导对细胞外酸化的影响,这与通过CO2的水合作用形成HCO3和H+的营养氧化作用相吻合,其程度高于FCCP3。然而,在分离的线粒体和透化细胞的情况下,如果以正确的浓度滴定,这些解耦合剂中的任何一个都应该与BAM15一样用于线粒体解偶联。
化合物渗透和测定循环的动力学
使用XF分析仪对3D球体进行典型 有丝体应力测试 的化合物的浓度、穿透力和动力学特征更难解决。鉴于球状体呈现3D结构,分子在球体直径上的穿透是一个比细胞单层复杂得多的过程。例如,在HepG2球体模型36中,动力学穿透性以及因此对化疗药物索拉非尼的敏感性是由球体年龄决定的,因此由大小决定。小分子化学物质(例如,药物,纳米颗粒)达到生物目标的能力取决于几个潜在的因素,包括通过37,38动态穿透和扩散的系统的复杂性。对于靶向肿瘤组织39的药物尤其如此。与3D球体背景下的肿瘤靶向类似,大小、致密性和其他表型反应(例如药物转运蛋白的表达)可以控制引发生物反应所需的化合物的渗透时间和浓度。
在该协议中,我们解决了ATP合酶抑制剂寡霉素,质子团和线粒体解偶联剂BAM15以及复合物I和复合III抑制剂鱼藤酮和抗霉素A的组合的穿透时间和小分子浓度问题。通过探测暴露于这些常见呼吸化合物的多次滴定的单MCF-7球体的OCR,我们证明诱导稳态呼吸频率所需的每种化合物的最佳浓度落在与单层细胞相似的范围内(图4)。重要的是,与单层对应物不同,结果表明,增加进样之间的测量周期数是在单个3D球体中实现稳态OCR的关键。这些数据强调了使用这些方法探索3D球体的线粒体呼吸参数时化合物渗透的重要性及其各自的动力学特征。使用由 图3, 图4和图 5中提供的数据告知的球体优化特性,化合物浓度和测量周期时间,建立了经过验证的 MitoStress测试 ,用于探测一系列癌症衍生的3D球体中线粒体氧化代谢的特定参数(图6)。重要的是,像一些单层癌细胞系40一样,某些癌症衍生的3D球体的最大呼吸能力(非耦合刺激呼吸速率)被寡霉素抑制(图5)。具体而言,与没有寡霉素的BAM15解偶联相比,从A549或MCF-7细胞生长的3D球体在寡霉素注射后与BAM15解偶联时显示出显着较低的最大呼吸速率(图5F 和 图5H)。鉴于这种效应可能存在于其他3D球体培养物中,我们建议除非采用先前验证的方案,否则应在不使用寡霉素的情况下估计3D球体的最大呼吸能力。
同时收集ECAR数据,作为细胞球体中糖酵解通量的量度
正如技术制造商的文献或信息中通常看到的那样,以ECAR形式测量的球体的糖酵解速率是可以与OCR一起捕获的次要参数。在任何XF实验中,单独计算ECAR都不是有用或有意义的参数,因为它没有针对XF测定缓冲液的缓冲能力或线粒体酸化的添加进行校正,线粒体酸化是由CO2 到HCO3- 和H +的水合引起的。ECAR只有在应用这些数据校正后才有见地,之后就可以提供有关糖酵解通量的更准确结论。为了校正缓冲能力以生成更有意义的PER数据,必须知道球状微孔板的微室体积。制造商无法用球状微孔板为此提供真实体积,因此,PER数据无法轻易确定。事实上,尽管这些测量可以通过经验来实现,但这超出了本手稿的范围。然而,通过适当的校正并知道孔中存在的给定球体尺寸的微室体积(例如,获得球体密度的测量值),ECAR数据将变得有意义,并且可以进行糖酵解PER的计算。因此,XF数据可以为研究球体中的糖酵解和氧化代谢提供更多信息,但前提是要深入考虑这些参数。
球体形成、处理、转移和移动
一些细胞系比其他细胞系更适合形成球状体,并且可能根本不形成球状体,例如,与其他细胞系相比,MCF-7卵巢癌细胞41,42 形成高度圆形的球状体(图3)。作为另一个例子,Capan-1胰腺癌细胞已被证明可以形成比Panc-1或BxPC343更好的球状体。同样,已知肝癌细胞系具有形成致密球体5,44的可变能力,观察到表型的变化,例如药物代谢增强或白蛋白的产生,例如HepG2与HepG2 / C3A9,45,46 或HepaRG球体17,47,48.因此,用户应相应地优化球体培养技术并进行滴定实验,以确定最佳的接种密度和培养时间过程。此外,测定培养基的制剂和组合物已被证明会影响球体制剂,包括甲基纤维素的加入,通常加入到培养基中以增加基质粘度43,49,50。因此,应根据经验确定所有使用的细胞系的最佳细胞培养基组成。
整个球体培养基交换的次数由所使用的细胞系决定。然而,通常,每2-3天进行一次半体积的培养基交换适用于大多数情况下补充营养。我们使用强制聚合方法使用来自市售来源的细胞排斥微孔板生成3D球体,以便在XF分析研究中快速开发和部署球体模型。然而,替代平台可能更适合从其他细胞类型生成球状体,例如,悬落或基质嵌入方法。在资源有限的实验室中,用户可能希望参考琼脂糖-液体覆盖技术来形成驱细胞微孔板表面51,52 以显著降低初始球状体方法开发步骤的经济成本。培养皿之间的球体移动对于进行XF分析和其他下游测定是必要的。转移的难易程度通常由球体尺寸和总密度决定。我们建议使用P200或P1000宽孔移液器吸头以保持球体完整性;小口径移液器吸头有机械破坏球体的风险,球状体可以通过商业采购购买,或者小心地通过简单地修剪移液器吸头的末端以增加孔口来制造。然而,这种方法可能会在尖端末端周围的塑料上引入毛皮,这可能导致处理过程中的机械破坏。背光灯或灯箱的使用对于在解剖显微镜下处理和观察球体也很有用,这是确保将球体成功转移到球体测定微孔板中的重要步骤。此外,在典型的有 丝分裂激素测试 中,球体测定微孔板内的球体位置特别重要,并直接影响OCR和复合效应(图7),这很可能是由于球体位置与传感器探针荧光团的关系。
背景校正和温度控制井
使用基于微孔板的测定是几个研究领域广泛使用的方法;然而,它们的使用带来了一些实际挑战。与其他实验方法一样,特别是那些使用96(或更大)阵列格式的方法,微孔板的几何形状和定位会随着时间的推移影响整个板的温度和气体交换梯度,通常称为“边缘效应”53,54。我们发现球体测定微孔板也是如此。根据制造商的指南和协议,最外层的角孔:A1、A12、H1 和 H12 始终被指定为 XFe96 分析仪的背景校正和温度控制孔。相反,对于24孔阵列格式,A1和D6被指定为对照孔,另外两个孔均匀分布在B4和C3的板中间。在执行XF球体分析时,我们发现最初使用制造商指南收集的数据存在显着偏差。尽管在开始采集数据之前,包括了必要的步骤来确保测定预平衡到温度和CO2 含量,但在注射某些呼吸抑制剂后,OCR通常产生负值(图8)。
我们发现这些观察结果可能是由于球体测定微孔板的边缘效应。在 图8中,我们发现在微孔板上重新分配背景控制孔,XF数据被调整了大约2倍。两个最可能的原因是(1)由于边缘孔的蒸发效应导致XFe96探针取样的总体积室较小,以及(2)指定用于背景校正的孔和样品孔之间的温度平衡不足,导致数据集掩盖或过度膨胀OCR。因此,为了避免这种结果,建议用户在整个球体测定微孔板中重新分配指定用于背景校正的孔,并采取必要步骤在获取XF数据之前预先平衡其测定。
数据的规范化
除了提供使用XF技术探测单个3D球体的线粒体能量代谢的详细方案外,本文还提出了将3D球体获得的线粒体呼吸频率数据归一化的可能方法。使用在不同细胞接种密度下培养的MCF-7球体获得的呼吸频率数据(图3),我们呈现了从MCF-7球体的基础线粒体呼吸速率,当归一化到初始细胞接种密度,球体体积和dsDNA含量时,其大小和直径增加(图9)。适当的归一化方法对于XF数据集的准确解释至关重要,特别是在比较 体外 3D球体模型和不同细胞类型时。规范化不良可能导致错误的结果,根本无法在数据集之间进行比较。蛋白质含量不是球体XF数据归一化的首选,因为预处理可能会影响蛋白质合成速率,而不会对呼吸频率产生显着影响。此外,大量不一致的蛋白质可以在细胞裂解时与球状微孔板结合,从而在孔之间引入蛋白质含量的变化。在使用球状体或非粘附细胞的XF分析中,这可能会进一步复杂化,这些细胞需要生物分子胶水来结合,其中可能含有蛋白质。
与细胞内蛋白质含量相反,核DNA含量与细胞类型无关,并且与细胞数量成正比(图9D)-比分离用于细胞数定量的球体更准确,更省时的方法。相反,Yepéz等人55在成纤维细胞的单层中进行XF分析,发现将XF数据归一化为细胞数量比归一化之前引入了更大的数据分散。核DNA含量与分化状态或表型无关,因此对于XF测定中球体数据的归一化比蛋白质含量更准确。DNA含量也已成为分析其他代谢相关数据集的成熟策略56。然而,重要的是要注意,核DNA含量是从球体中存在的所有细胞中定量的;因此,对于XF数据集,不建议对DNA含量进行归一化,其中球体接受可能导致细胞活力显着丧失的处理。对于此类数据集,如果可行,最好对细胞活力进行归一化,或者可以将数据基线校正为基础呼吸。
使用备用呼吸能力作为数据正常化重要性的示例
备用呼吸能力是最大线粒体呼吸能力减去基础线粒体呼吸频率的量度(图6)。然而,将这种类型的数据报告为速率(即某些实验中的pmolO2 / min / well)的问题在于数据没有归一化。即使将球状体数据归一化为细胞密度/DNA含量,这通常也排除了细胞内线粒体密度需要归一化的关键参数。鉴于线粒体密度的变化将导致基础呼吸和最大呼吸成比例的变化,备用容量也将增加。例如,如果球体 OCR基础 为 200,OCR最大值 为 400,则报告为 200;如果 OCR基础 为 100 且 OCR最大值,则备用容量也为 100;然而,作为一个百分比,它们都是最大值的50%(或基础的100%)。因此,尽管以 pmols O 2/min/well 计算时,速率为200 和 100,但备用容量在这两个示例之间没有变化。内部归一化的值更可靠和更有见地,使XF数据在研究和项目中更具可比性。为了节省呼吸能力,我们选择将其表示为最大呼吸的百分比,而不是绝对速率。这也可以表示为基础呼吸的百分比。如果使用细胞或球体,情况就是如此。然而,鉴于微孔板中球体的位置可能会改变绝对OCR,但不会改变抑制剂或解耦合器的相对变化,因此更重要的是将球体中的内部归一化反应视为折叠变化或百分比。
这里生成的球体模型呈现出一系列经典2D模型中无法捕获的细胞类型和结构。这些包括异质的、三维细胞的空间排列、增强的细胞-细胞接触(例如,间隙连接和细胞外基质的形成)以及整个球体直径的生化梯度(例如,pH梯度、氧扩散对营养物质的获取)。使用细胞外通量研究 体外 球体生物学可以通过代谢扰动观察来确定药物治疗的最佳靶点。这些可以从 体外 球体外推到 体内 肿瘤,并确定可能靶向球状肿瘤代谢的途径,例如,球体生长期间的碳水化合物利用。治疗方式在早期生长阶段靶向球体方面可能有效,但随着代谢网络复杂性的成熟,在球状体生长的后期阶段被证明效果较差。总而言之,3D细胞培养模型和复杂分析技术在生物学研究中的发展将继续是一个充满活力和快速变化的领域,具有无与伦比的潜力。 体外 细胞培养球体的细胞外通量分析可以作为一种前沿研究方法,以推进可以外推的研究结果,以更好地理解人类相关生物学,减少研究中动物模型的使用,并加强以患者为中心的研究。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
作者没有利益冲突需要声明。
Acknowledgments
新泽西州获得了与系统发现有限公司(BB/M01116X/1,1940003)的BBSRC MIBTP案例奖的支持
Materials
| Name | Company | Catalog Number | Comments |
| A549 | ECACC | #86012804 | Lung carcinoma cell line |
| Agilent Seahorse XF RPMI Medium, pH 7.4 | Agilent Technologies Inc. | 103576-100 | XF assay medium with 1 mM HEPES, without phenol red, sodium bicarbonate, glucose, L-glutamine, and sodium pyruvate |
| Agilent Seahorse XFe96 Extracellular Flux Analyzer | Agilent Technologies Inc. | - | Instrument for measuring rates of spheroid oxygen uptake in single spheroids |
| Antimycin A | Merck Life Science | A8674 | Mitochondrial respiratory complex III inhibitor |
| BAM15 | TOCRIS bio-techne | 5737 | Mitochondrial protnophore uncoupler |
| Black-walled microplate | Greiner Bio-One | 655076 | For fluorescence-based assays |
| CELLSTAR cell-repellent surface 96 U well microplates | Greiner Bio-One | 650970 | Microplates for generating spheroids |
| CellTiter-Glo 3D Cell Viability Assay | Promega | G9681 | Assay for the determination of cell viability in 3D microtissue spheroids |
| Cultrex Poly-D-Lysine | R&D Systems a biotechne brand | 3439-100-01 | Molecular cell adhesive for coating XFe96 spheroid microplates to facillitate attachment of spheroids |
| D-(+)-Glucose | Merck Life Sciences | G8270 | Supplement for cell culture growth and XF assay medium |
| Dulbecco’s Modified Eagle Medium (DMEM) | Gibco | 11885084 | Culture medium for HepG2/C3A spheroids |
| EVOS XL Core Imaging System | Thermo Fisher Scientific | AMEX1000 | Phase-contrast imaging microscope |
| EZ-PCR Mycoplasma test kit | Biological Industries | 20-700-20 | Mycoplasma screening in cell cultures |
| FIJI Is Just Image J | Analysis of collated images | ||
| Foetal bovine serum | Merck Life Science | F7524 | Supplement for cell culture medium |
| HepG2/C3A | ATCC | #CRL-10741 | Hepatic carcinoma cell line, a clonal derivative of the parent HepG2 cell line |
| Lactate-Glo | Promega | J5021 | Assay for measurement of lactate within spheorid culture medium |
| L-glutamine (200 mM solution) | Merk Life Sciences | G7513 | Supplement for cell culture growth and XF assay medium |
| M50 Stereo microscope | Leica Microsytems | LEICAM50 | Stereo dissection micrscope; used for spheorid handling |
| MCF-7 | ECACC | #86012803 | Breast adenocarcinoma cell line |
| Oligomycin from Streptomyces diastatochromogenes | Merck Life Science | O4876 | ATP Synthase Inhibitor |
| Penicilin-Streptomycin | Gibco | 15140122 | Antibiotics added to cell culture medium |
| Quant-iT PicoGreen dsDNA Assay Kit | Initrogen | P7589 | Analysis of dsDNA in spehroids |
| Rotenone | Merck Life Science | R8875 | Mitochondrial Respiratory Complex I Inhibitor |
| RPMI 1640 | Gibco | 21875091 | Culture medium for A549, MCF7, and SK-OV-3 spheroids |
| Seahorse Analytics | Agilent Technologies Inc. | Build 421 | https://seahorseanalytics.agilent.com |
| Seahorse XFe96 Spheroid FluxPak | Agilent Technologies Inc. | 102905-100 | Each Seahorse XFe96 Spheroid FluxPak contains: 6 Seahorse XFe96 Spheroid Microplates (102978-100), 6 XFe96 sensor cartridges, and 1 bottle of Seahorse XF Calibrant Solution 500 mL (100840-000) |
| Serological pipette: 5, 10, and 25 mL | Greiner Bio-One | 606107; 607107; 760107 | Consumables for cell culture |
| SK-OV-3 | ECACC | #HTB-77 | Ovarian adenocarcinoma cell line |
| Sodium pyruvate (100 mM solution) | Merck Life Science | S8636 | Supplement for cell culture growth and XF assay medium |
| T75 cm2 cell culture flask | Greiner Bio-One | 658175 | Tissue culture treated flasks for maintaining cell cultures |
| TrypLExpress | Gibco | 12604-021 | Cell dissociation reagent |
| Wave controller software | Agilent Technologies Inc. | - | |
| Wide orifice tip | STARLAB International GmbH | E1011-8400 | Pipette tips with wide opening for spheroid handling |
References
- Correa de Sampaio, P., et al. A heterogeneous in vitro three dimensional model of tumour-stroma interactions regulating sprouting angiogenesis. PLoS One. 7 (2), 30753 (2012).
- Amann, A., et al. Development of an innovative 3D cell culture system to study tumour-stroma interactions in non-small cell lung cancer cells. PLoS One. 9 (3), 92511 (2014).
- Russell, S., Wojtkowiak, J., Neilson, A., Gillies, R. J. Metabolic profiling of healthy and cancerous tissues in 2D and 3D. Scientific Reports. 7 (1), 15285 (2017).
- Zanoni, M., et al. 3D tumor spheroid models for in vitro therapeutic screening: a systematic approach to enhance the biological relevance of data obtained. Scientific Reports. 6, 19103 (2016).
- Song, Y., et al. Patient-derived multicellular tumor spheroids towards optimized treatment for patients with hepatocellular carcinoma. Journal of Experimental and Clinica Cancer Research. 37 (1), 109 (2018).
- Courau, T., et al. Cocultures of human colorectal tumor spheroids with immune cells reveal the therapeutic potential of MICA/B and NKG2A targeting for cancer treatment. Journal for ImmunoTherapy of Cancer. 7 (1), 74 (2019).
- Ivanova, E., et al. Use of ex vivo patient-derived tumor organotypic spheroids to identify combination therapies for HER2 mutant non-small cell lung cancer. Clinical Cancer Research. 26 (10), 2393-2403 (2020).
- Mandon, M., Huet, S., Dubreil, E., Fessard, V., Le Hegarat, L. Three-dimensional HepaRG spheroids as a liver model to study human genotoxicity in vitro with the single cell gel electrophoresis assay. Scientific Reports. 9 (1), 10548 (2019).
- Stampar, M., et al. Hepatocellular carcinoma (HepG2/C3A) cell-based 3D model for genotoxicity testing of chemicals. Science of the Total Environment. 755, 143255 (2020).
- Coltman, N. J., et al. Application of HepG2/C3A liver spheroids as a model system for genotoxicity studies. Toxicology Letters. 345, 34-45 (2021).
- Tchoryk, A., et al. Penetration and uptake of nanoparticles in 3D tumor spheroids. Bioconjugate Chemistry. 30 (5), 1371-1384 (2019).
- Leite, P. E. C., et al. Suitability of 3D human brain spheroid models to distinguish toxic effects of gold and poly-lactic acid nanoparticles to assess biocompatibility for brain drug delivery. Partical Fibre Toxicology. 16 (1), 22 (2019).
- Elje, E., et al. Hepato(Geno)toxicity assessment of nanoparticles in a HepG2 liver spheroid model. Nanomaterials. 10 (3), 545 (2020).
- Conway, G. E., et al. Adaptation of the in vitro micronucleus assay for genotoxicity testing using 3D liver models supporting longer-term exposure durations. Mutagenesis. 35 (4), 319-330 (2020).
- Wang, Z., et al. HepaRG culture in tethered spheroids as an in vitro three-dimensional model for drug safety screening. Journal of Applied Toxicology. 35 (8), 909-917 (2015).
- Proctor, W. R., et al. Utility of spherical human liver microtissues for prediction of clinical drug-induced liver injury. Archives of Toxicology. 91 (8), 2849-2863 (2017).
- Basharat, A., Rollison, H. E., Williams, D. P., Ivanov, D. P. HepG2 (C3A) spheroids show higher sensitivity compared to HepaRG spheroids for drug-induced liver injury (DILI). Toxicology and Applied Pharmacology. 408, 115279 (2020).
- Benning, L., Peintner, A., Finkenzeller, G., Peintner, L. Automated spheroid generation, drug application and efficacy screening using a deep learning classification: a feasibility study. Scientific Reports. 10 (1), 11071 (2020).
- Mittler, F., et al. High-content monitoring of drug effects in a 3D spheroid model. Frontiers in Oncology. 7, 293 (2017).
- Brand, M. D., Nicholls, D. G.
- Benz, R., McLaughlin, S. The molecular mechanism of action of the proton ionophore FCCP (carbonylcyanide p-trifluoromethoxyphenylhydrazone). Biophysical Journal. 41 (3), 381-398 (1983).
- Kasianowicz, J., Benz, R., McLaughlin, S. The kinetic mechanism by which CCCP (carbonyl cyanide m-chlorophenylhydrazone) transports protons across membranes. The Journal of Membrane Biology. 82 (2), 179-190 (1984).
- Kenwood, B. M., et al. Identification of a novel mitochondrial uncoupler that does not depolarize the plasma membrane. Molecular Metabolism. 3 (2), 114-123 (2013).
- Mitchell, P. Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature. 191, 144-148 (1961).
- Alexopoulos, S. J., et al. Mitochondrial uncoupler BAM15 reverses diet-induced obesity and insulin resistance in mice. Nature Communications. 11 (1), 2397 (2020).
- Chen, S. -Y., et al. Mitochondrial uncoupler SHC517 reverses obesity in mice without affecting food intake. Metabolism - Clinical and Experimental. 117, 154724 (2021).
- Goedeke, L., Shulman, G. I. Therapeutic potential of mitochondrial uncouplers for the treatment of metabolic associated fatty liver disease and NASH. Molecular Metabolism. 46, 101178 (2021).
- Hill, B. G., et al. Integration of cellular bioenergetics with mitochondrial quality control and autophagy. Biological chemistry. 393 (12), 1485-1512 (2012).
- Demine, S., Renard, P., Arnould, T. Mitochondrial uncoupling: a key controller of biological processes in physiology and diseases. Cells. 8 (8), 795 (2019).
- Wang, J., et al. Uncoupling effect of F16 is responsible for its mitochondrial toxicity and anticancer activity. Toxicological Sciences. 161 (2), 431-442 (2018).
- Tretter, L., Chinopoulos, C., Adam-Vizi, V. Plasma membrane depolarization and disturbed Na+ homeostasis induced by the protonophore carbonyl cyanide-p-trifluoromethoxyphenyl-hydrazon in isolated nerve terminals. Molecular Pharmacology. 53 (4), 734-741 (1998).
- Connop, B. P., Thies, R. L., Beyreuther, K., Ida, N., Reiner, P. B. Novel effects of FCCP [carbonyl cyanide p-(trifluoromethoxy) phenylhydrazone] on amyloid precursor protein processing. Journal of neurochemistry. 72 (4), 1457-1465 (1999).
- Stöckl, P., et al. Partial uncoupling of oxidative phosphorylation induces premature senescence in human fibroblasts and yeast mother cells. Free Radical Biology and Medicine. 43 (6), 947-958 (2007).
- Firsov, A. M., et al. Protonophoric action of BAM15 on planar bilayers, liposomes, mitochondria, bacteria and neurons. Bioelectrochemistry. 137, 107673 (2021).
- Dranka, B. P., Hill, B. G., Darley-Usmar, V. M. Mitochondrial reserve capacity in endothelial cells: The impact of nitric oxide and reactive oxygen species. Free Radical Biology and Medicine. 48 (7), 905-914 (2010).
- Eilenberger, C., Rothbauer, M., Ehmoser, E. K., Ertl, P., Kupcu, S. Effect of spheroidal age on sorafenib diffusivity and toxicity in a 3D HepG2 spheroid model. Scientific Reports. 9 (1), 4863 (2019).
- vanden Brand, D., Veelken, C., Massuger, L., Brock, R. Penetration in 3D tumor spheroids and explants: Adding a further dimension to the structure-activity relationship of cell-penetrating peptides. Biochimica et Biophysica Acta (BBA) - Biomembranes. 1860 (6), 1342-1349 (2018).
- Niora, M., et al. Head-to-head comparison of the penetration efficiency of lipid-based nanoparticles into tumor spheroids. ACS Omega. 5 (33), 21162-21171 (2020).
- Millard, M., et al. Drug delivery to solid tumors: the predictive value of the multicellular tumor spheroid model for nanomedicine screening. International Journal of Nanomedicine. 12, 7993-8007 (2017).
- Ruas, J. S., et al. Underestimation of the maximal capacity of the mitochondrial electron transport system in oligomycin-treated cells. PLoS One. 11 (3), 0150967 (2016).
- Benton, G., DeGray, G., Kleinman, H. K., George, J., Arnaoutova, I. In vitro microtumors provide a physiologically predictive tool for breast cancer therapeutic screening. PLoS One. 10 (4), 0123312 (2015).
- Hirpara, J., et al. Metabolic reprogramming of oncogene-addicted cancer cells to OXPHOS as a mechanism of drug resistance. Redox Biology. 25, 101076 (2019).
- Ware, M. J., et al. Generation of homogenous three-dimensional pancreatic cancer cell spheroids using an improved hanging drop technique. Tissue Engineering. Part C, Methods. 22 (4), 312-321 (2016).
- Song, Y., et al. TGF-β-independent CTGF induction regulates cell adhesion mediated drug resistance by increasing collagen I in HCC. Oncotarget. 8 (13), 21650-21662 (2017).
- Wrzesinski, K., et al. HepG2/C3A 3D spheroids exhibit stable physiological functionality for at least 24 days after recovering from trypsinisation. Toxicology Research. 2 (3), 163-172 (2013).
- Gaskell, H., et al. Characterization of a functional C3A liver spheroid model. Toxicology Research. 5 (4), 1053-1065 (2016).
- Takahashi, Y., et al. 3D spheroid cultures improve the metabolic gene expression profiles of HepaRG cells. Bioscience Reports. 35 (3), 00208 (2015).
- Hendriks, D. F. G., Puigvert, L. F., Messner, S., Mortiz, W., Ingelman-Sundberg, M. Hepatic 3D spheroid models for the detection and study of compounds with cholestatic liability. Scientific Reports. 6, 35434 (2016).
- Leung, B. M., Lesher-Perez, S. C., Matsuoka, T., Moraes, C., Takayama, S. Media additives to promote spheroid circularity and compactness in hanging drop platform. Biomaterials Science. 3 (2), 336-344 (2015).
- Cavo, M., et al. A synergic approach to enhance long-term culture and manipulation of MiaPaCa-2 pancreatic cancer spheroids. Scientific Reports. 10 (1), 10192 (2020).
- Carlsson, J., Yuhas, J. M.
- Costa, E. C., Gaspar, V. M., Coutinho, P., Correia, I. J. Optimization of liquid overlay technique to formulate heterogenic 3D co-cultures models. Biotechnology and Bioengineering. 111 (8), 1672-1685 (2014).
- Lundholt, B. K., Scudder, K. M., Pagliaro, L. A simple technique for reducing edge effect in cell-based assays. Journal of Biomolecular Screening. 8 (5), 566-570 (2003).
- Zhang, X. D., et al. The use of strictly standardized mean difference for hit selection in primary RNA interference high-throughput screening experiments. Journal of Biomolecular Screening. 12 (4), 497-509 (2007).
- Yepez, V. A., et al. OCR-Stats: Robust estimation and statistical testing of mitochondrial respiration activities using Seahorse XF Analyzer. PLoS One. 13 (7), 0199938 (2018).
- Silva, L. P., et al. Measurement of DNA concentration as a normalization strategy for metabolomic data from adherent cell lines. Analytical Chemistry. 85 (20), 9536-9542 (2013).
Tags
癌症研究,第180期,Erratum
Formal Correction: Erratum: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis
Posted by JoVE Editors on 03/11/2022.
Citeable Link.
An erratum was issued for: Exploring Mitochondrial Energy Metabolism of Single 3D Microtissue Spheroids using Extracellular Flux Analysis. The Representative Results section was updated.
Figure 5 was updated from:
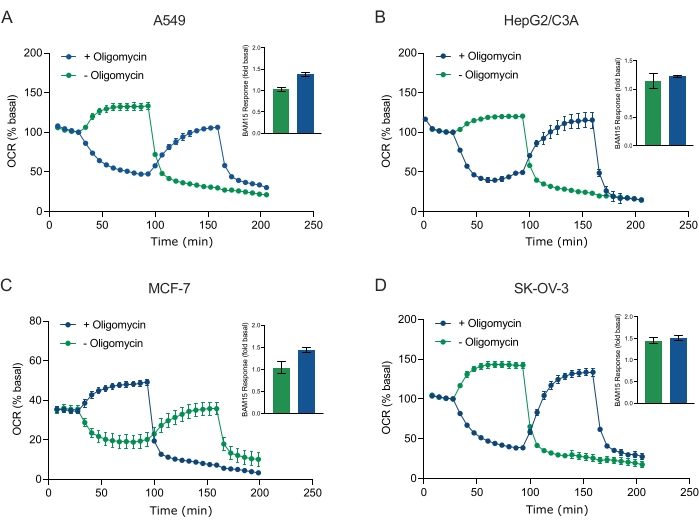
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
to:
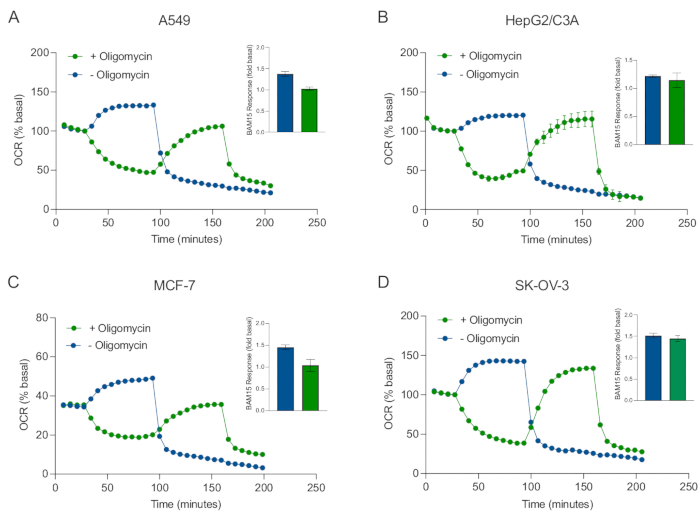
Figure 5: Single or sequential injection of mitochondrial respiratory compounds. Cancer-cell-derived spheroids of MCF-7, HEPG2/C3A, SK-OV-3, and A549 were placed into wells of an XFe96 spheroid microplate in XF RPMI and probed for OCR using the Agilent Seahorse XFe96 analyzer. OCR was measured 5x, after which 2 µg/mL oligomycin (injection Port A: green trace) or 5 µM BAM15 (injection Port A: blue trace or injection port B: green trace) to inhibit the mitochondrial ATP synthase and determine maximal respiratory capacity, respectively. Kinetic OCR data are expressed as % basal (A-D). Maximal respiratory capacity (OCRmax) was calculated as a factor of basal OCR by the equation: OCRmax = OCRBAM15 / OCRbasal. OCRmax was obtained from OCR averages across measurement cycles 8-10 post BAM15 injection with (green bars) and without (blue bars) oligomycin. Data are averages ± SEM from 3-8 individual well replicates across the spheroid assay microplate. Abbreviations: OCR = oxygen consumption rate. Please click here to view a larger version of this figure.
