Ved at levere og fjerne proteiner til og fra plasmamembranen styrer membranproteinhandel i neuroner plasmamembranudstyret med receptorer såvel som ionkanaler og regulerer som følge heraf neuronal funktion. Misregulering eller defekter i proteinhandel har typisk skadelige virkninger på celler og resulterer i neuronal degeneration. Hos mennesker kan dette forårsage neurodegenerative sygdomme som Alzheimers og Parkinsons sygdom eller Retinitis pigmentosa1. Fotoreceptorer i drosophila melanogasters sammensatte øje er blevet et in vivo-modelsystem til undersøgelse af membranproteinhandel2. Dette skyldes ikke kun Drosophilas genetiske alsidighed, der muliggør effektive genetiske skærme, men også fordi alle væsentlige komponenter i den lysabsorberende fotoreceptormembran er karakteriseret i detaljer, og der findes effektive mikroskopiske teknikker, der kan påføres flueøjet. Disse teknikker er i fokus i denne artikel.
I Drosophila fotoreceptorceller danner den apikale plasmamembran en tæt pakket stak mikrovilli langs den ene side af cellen, nævnt rhabdomere. Rhabdomeres af fotoreceptorceller R1-6 er arrangeret i et karakteristisk trapezformet mønster, mens fotoreceptorcellerne R7 og R8 danner en enkelt rhabdomere i midten af denne trapez3. Membranproteinhandel er nødvendig for en reguleret omsætning af rhabdomerale membranproteiner såsom rhodopsin og de lysaktiverede TRP (forbigående receptorpotentiale) og TRPL (TRP-lignende) ionkanaler for at sikre den korrekte mængde af disse fototransduktionsproteiner i rhabdomere. Fotoreceptormembranproteiner syntetiseres i det endoplasmatiske retikulum og transporteres via Golgi-apparatet til rhabdomere. Efter aktivering af rhodopsin ved lys kan et rhodopsinmolekyle enten blive inaktiveret ved absorption af en anden foton eller kan fjernes fra rhabdomere ved clathrinmedieret endocytose. Endocytosed rhodopsin bliver enten nedbrudt i lysosomet eller genbruges tilbage til rhabdomere 4,5. Ionkanalen TRPL internaliseres også efter aktivering af fototransduktionskaskaden og gennemgår en lysafhængig translokation mellem rhabdomere (hvor den er placeret, når fluer holdes i mørket) og et ER-beriget opbevaringsrum i cellelegemet (hvortil det transporteres inden for flere timer ved belysning)6,7,8,9,10 . I modsætning til endocytoseret rhodopsin nedbrydes kun små mængder TRPL via den endolysosomale vej, og størstedelen opbevares intracellulært i stedet og genbruges tilbage til rhabdomere ved mørk tilpasning6. TRPL kan således bruges til at analysere lysudløst handel med plasmamembranproteiner. Drosophila fotoreceptorceller anvendes også til at studere neuronal degeneration. Fotoreceptorcelledegeneration bestemmes ofte ved at vurdere strukturen af rhabdomeres, som opløses som følge af degenerative processer5.
For at studere den subcellulære lokalisering af TRPL og rhodopsin i fotoreceptorceller eller fotoreceptorcelledegeneration er der anvendt to fluorescensmikroskopimetoder, der adskiller sig med hensyn til analysehastighed og opløsning. En meget hurtig, ikke-invasiv metode, der kan bruges til genetiske skærme, men med en begrænset rumlig opløsning, er påvisning af fluorescens i den dybe pseudopupil (DPP). DPP er et optisk fænomen af leddyr sammensatte øjne, hvis geometriske oprindelse er blevet forklaret detaljeret af Franceschini og Kirschfeld i 197111. Kort sagt, på flere optiske planer under nethinden kan overlay-billeder af rhabdomeres fra tilstødende ommatidia observeres. På et brændplan gennem midten af øjets krumning danner disse overlejrede fremspring et billede, der ligner det trapezformede layout af rhabdomeres i et enkelt ommatidium, kun størrelsesordener større. Dette fænomen kan også observeres uafhængigt af eksogen ekspression af fluorescensproteiner (f.eks. TRPL::eGFP8), hvilket ikke desto mindre gør DPP lettere at detektere (figur 1A-A”)12. En anden ikke-invasiv metode er vandnedsænkningsmikroskopi, der er afhængig af billeddannelse af fluorescerende mærkede proteiner efter optisk neutralisering af øjnenes dioptriske apparat med vand (figur 1B-C”)12. Ved hjælp af vandnedsænkningsmetoden kan den relative mængde TRPL::eGFP i rhabdomeres eller cellelegemet vurderes kvantitativt for individuelle fotoreceptorceller. Desuden kan ikke-translokerende fluorescensmærkede proteiner anvendes til at evaluere rhabdomeral integritet og til at bestemme tidsforløbet for en potentiel degeneration på en kvantitativ måde, som beskrevet her.
Mens optagelser af DPP er langt den nemmeste og hurtigste af disse metoder at udføre, er den rumlige opløsning af data, de genererer, begrænset. Derudover er der mange grunde til, at en DPP kan være fraværende, som ikke nødvendigvis kan ses af DPP-billeddannelse selv. Da DPP repræsenterer en summation af flere ommatidia, går information om individuelle celler tabt. DPP-billeddannelse med lav opløsning tjener således en vigtig funktion ved screening af et stort antal fluer, men bør generelt efterfølges af optagelser med højere opløsning ved hjælp af vandnedsænkningsmikroskopi. Vandnedsænkningsmikrografer tillader fortolkninger om individuelle celler, udviklingsdefekter, øjenmorfologi, proteinfejllokalisering eller retinal degeneration samt kvantificering af disse effekter. Denne protokol beskriver disse to teknikker i detaljer.
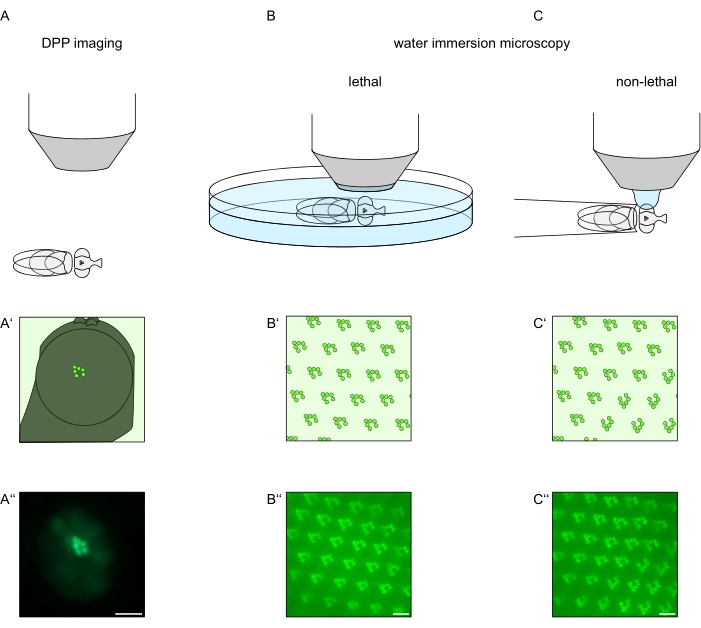
Figur 1: Oversigt over mikroskopivariationer for Drosophila-øjet, der præsenteres i denne protokol. Skematiske repræsentationer og eksemplariske mikrografer af (A-A”) fluorescerende dyb pseudopupil (DPP) billeddannelse, (B-B”) dødelig vandnedsænkningsmikroskopi af fluorescerende rhabdomere og (C-C”) ikke-dødelig vanddråbemikroskopi af fluorescerende rhabdomere. Skalabjælke (A”): 100 μm. Vægtstænger (B”-C”): 10 μm. Figuren er ændret i forhold til reference13. Klik her for at se en større version af denne figur.
