The first step of this study was to optimize the slice thickness and punch size used to excise a section of the striatum from the slice (Figure 3A). A slice at 150 µm thickness and a 1.5 mm punch size gave the best results determined by the coupling efficiency (Figure 3B–C). As shown in Figure 3B, OCR is relatively stable for 5 h with less than 10% run down. In addition, functional measurements were used, as well as patch-clamp recording of cortical neurons and medium spiny neurons in the striatum, and fast-scan cyclic voltammetry (FSCV) measurement of DA release, to demonstrate that the neurons and terminals were fully functional in this preparation, as shown in Zhi et al.24. Included with this, we then tested different concentrations of FCCP to discover which would give the best response; 10 µM FCCP gave the best readout (Figure 3D1,D2) determined by the spare respiratory capacity:
Spare respiratory capacity = maximal respiration – basal respiration
These slice conditions were used to measure the differences in the OCR of striatal slices from PINK1 KO mice and wild-type (WT) littermates. For the results of PINK1 KO and WT mice, 3-4 mice were used and 4-6 replicates per mouse were used and averaged to obtain one data point.Since PINK1 is a key regulator of mitochondrial function, mitochondrial dysfunction was expected to be found in the KO mice, measured by decreased mitochondrial OCR. Indeed, an age-dependent decrease in OCR was evident in the KO mice compared to their WT controls (Figure 4A,D). Basal OCR was similar in both the KO and WT groups for young mice (3-4 months); however, basal OCR decreased for the KO mice in the old group (10-14 months; Figure 4B,E). Coupling efficiency, determined as below,
Coupling efficiency = [ATP production rate] / [basal respiration rate] × 100
however, decreased in the KO mice for both the young and old groups (Figure 4C,F). This data demonstrates that the sections of striatal tissue from PINK1 KO mice had mitochondrial dysfunction. This dysfunction in the KO mice also started from a young age, indicated by the decreased coupling efficiency, although the basal OCR was only decreased in the old group. This age-dependent decrease was likely a result of accumulating mitochondrial defects caused by the knockout of PINK1.
Figure 1: Images showing the extracellular flux analyzer hydrate cartridge sensors and plate. (A) The sensor cartridge sitting on top of a calibration plate (utility plate). (B) Side view of the utility plate and sensor cartridge plate. The triangular notch of the utility and sensor cartridge plate should be correctly aligned. Please click here to view a larger version of this figure.
Figure 2: Punched slice attachment and placement. (A) Punched slice is attached to the mesh insert of the capture screen, and then (B) placed into one of the wells of the incubating islet plate carefully to ensure the slice does not detach or move during the measurement. Please click here to view a larger version of this figure.
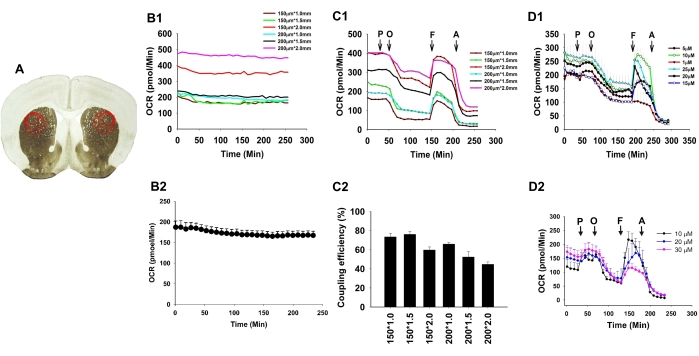
Figure 3: Optimal striatal slice condition for flux respiration assay. (A) Diagram of tissue punch sizes (1.0 mm, 1.5 mm, and 2.0 mm in diameter) for the striatum (STR) with red circles representing the areas that were obtained for the analysis. (B1) O2 consumption rates (OCRs) for different thicknesses and punch sizes of slices in the control group showed stable basal respiration over the whole measurement (4 h), and OCR was proportional to the volume of the slice. OCRs were measured in 150 µm and 200 µm thickness slices punched by 1.0 mm, 1.5 mm, and 2.0 mm punch. The combinations used are mentioned in the legends as thickness * punch size (e.g., 150 µm * 1 mm). (B2) Averaged OCRs measured in 150 µm thickness slices punched by 1.5 mm punch; n = 7. (C1) Representative OCR responses of slices at different thicknesses and diameters with 10 mM pyruvate (P), 20 µM oligomycin (O), 10 µM FCCP (F), and 20 µM antimycin A (A) injected sequentially. The arrows indicate the time point of injection of these compounds. (C2) The mitochondrial coupling efficiency was compared among different groups. Slices of 150 µm * 1.5 mm had the high coupling efficiency but the smallest variance; n = 4. (D1) Titration experiment for FCCP was performed, and 10 µM FCCP gave the biggest spare capacity; n = 4 for each group. (D2) Titration experiments for FCCP were performed with an analyzer, and 10 µM FCCP gave the biggest spare capacity; n = 4 for each group. The values given in the figures are mean ± standard error of the mean (SEM). This figure has been modified from Zhi et al.24. Please click here to view a larger version of this figure.
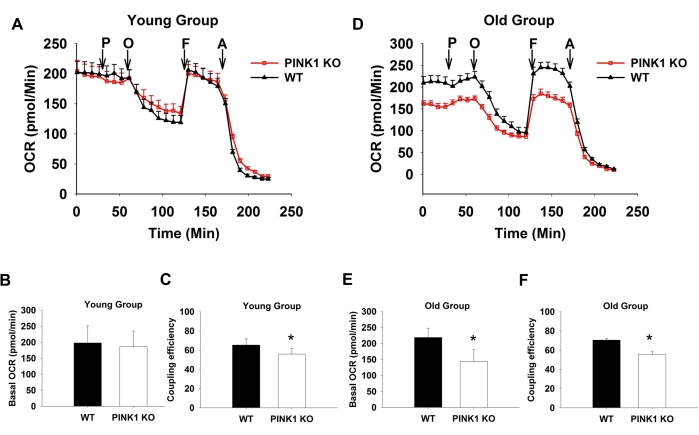
Figure 4: PINK1 KO slices from the old group showing significantly lower OCR. OCRs of acute STR slices (150 µm * 1.5 mm) from mice in the young (A, B, and C) and the old groups (D, E, and F), exposed to successive additions of respiratory modulators (showed using arrows). The OCR of the young group was not significantly different between different genotypes (B), while the coupling efficiency (determined as a percentage value using the formula mentioned above %) was decreased in PINK1 KO slices (C). In the old group, the PINK1 KOslices showed significantly decreased basal respiration level (E) and coupling efficiency (F). The following solution, 10 mM pyruvate (P), 20 µM oligomycin (O), 10 µM FCCP (F), and 20 µM antimycin A (A), was injected sequentially. n = 4 for each genotype and age. The values given in the figures are mean ± SEM. The difference was considered significant when p < 0.05 (*). This figure has been modified from Zhi et al.24. Please click here to view a larger version of this figure.
