Biomolekulare Wechselwirkungen:
Proteine sind wesentliche Bestandteile von Organismen und an zahlreichen molekularen Signalwegen wie Zellstoffwechsel, Zellstruktur, Zellsignalisierung, Immunantwort, Zelladhäsion und mehr beteiligt. Während einige Proteine ihre Funktion(en) unabhängig voneinander ausführen, interagieren die meisten Proteine mit anderen Proteinen über eine Bindungsgrenzfläche, um die richtige biologische Aktivität zu koordinieren1.
Biomolekulare Wechselwirkungen können hauptsächlich auf der Grundlage der unterschiedlichen strukturellen und funktionellen Eigenschaften der beteiligten Proteineklassifiziert werden 2, beispielsweise basierend auf den Proteinoberflächen, der komplexen Stabilität oder der Persistenz von Wechselwirkungen3. Die Identifizierung essentieller Proteine und ihrer Rolle in biomolekularen Wechselwirkungen ist entscheidend für das Verständnis biochemischer Mechanismen auf molekularer Ebene4. Derzeit gibt es verschiedene Ansätze, um diese Wechselwirkungen5 zu erkennen: in vitro 6, in silico7, in lebenden Zellen8, ex vivo 9 und in vivo10, wobei jede ihre eigenen Stärken und Schwächen hat.
Die In-vivo-Assays werden unter Verwendung des gesamten Tieres als experimentelles Werkzeugdurchgeführt 11, unddie Ex-vivo-Assays werden an Gewebeextrakten oder ganzen Organen (z. B. Herz, Gehirn, Leber) in einer kontrollierten äußeren Umgebung durchgeführt, wobei die natürlichen Bedingungen nur minimal verändert werden. Die häufigste Anwendung von In-vivo- und Ex-vivo-Studien besteht darin, die pharmakokinetischen, pharmakodynamischen und toxischen Wirkungen potenzieller pharmakologischer Wirkstoffe vor Studien am Menschen zu bewerten, indem ihre allgemeine Sicherheit und Wirksamkeit sichergestelltwird 12.
Biomolekulare Wechselwirkungen können auch innerhalb lebender Zellen nachgewiesen werden. Die Bildgebung lebender Zellen ermöglicht es uns, dynamische Wechselwirkungen zu beobachten, während sie die Reaktionen eines bestimmten biochemischen Weges ausführen13. Darüber hinaus können Detektionstechniken wie Biolumineszenz oder Fluoreszenzresonanz-Energietransfer Informationen darüber liefern, wo und wann diese Wechselwirkungen innerhalb der Zelle auftreten14. Obwohl der Nachweis in lebenden Zellen entscheidende Details bietet, stützen sich diese Nachweismethoden auf Optik und Markierungen, die möglicherweise nicht die native Biologie widerspiegeln. Sie sind auch weniger kontrolliert als In-vitro-Methoden und erfordern spezielles Fachwissen, um15 durchzuführen.
Die In-silico-Berechnungsmethoden werden in erster Linie für das groß angelegte Screening von Zielmolekülen vor den In-vitro-Experimenten eingesetzt. Computergestützte Vorhersagemethoden, computergestützte Datenbanken, molekulares Docking, quantitative Struktur-Aktivitäts-Beziehungen und andere molekulardynamische Simulationsansätze gehören zu den etablierten In-silico-Werkzeugen 16. Im Vergleich zu mühsamen experimentellen Techniken können die In-silico-Werkzeuge leicht Vorhersagen mit hoher Empfindlichkeit, aber mit reduzierter Genauigkeit in der Vorhersageleistungtreffen 17.
In-vitro-Assays werden mit Mikroorganismen oder biologischen Molekülen außerhalb ihres biologischen Standardkontexts durchgeführt. Die Darstellung biomolekularer Interaktionen durch In-vitro-Methoden ist entscheidend für das Verständnis von Proteinfunktionen und der Biologie hinter dem komplexen Netzwerk der Zellfunktion. Die bevorzugte Assay-Methodik wird entsprechend den intrinsischen Eigenschaften des Proteins, den kinetischen Werten und dem Modus und der Intensität der Wechselwirkungenausgewählt 18,19.
Die Hsp90/Cdc37-Interaktion:
Der Chaperon-Kinase-Signalweg, der Hsp90 und Cdc37 verbindet, ist ein vielversprechendes therapeutisches Ziel in der Tumorbiologie20. Hsp90 spielt eine zentrale Rolle bei der Kontrolle des Zellzyklus, der Proteinzusammenordnung, dem Zellüberleben und den Signalwegen. Proteine, die für ihre Funktionen auf Hsp90 angewiesen sind, werden zur Komplexierung durch ein Co-Chaperon wie Cdc37 an Hsp90 abgegeben. Der Hsp90/Cdc37-Komplex steuert die Faltung der meisten Proteinkinasen und dient als Drehscheibe für eine Vielzahl von intrazellulären Signalnetzwerken21. Es ist ein vielversprechendes Anti-Tumor-Ziel aufgrund seiner erhöhten Expression bei verschiedenen Malignomen, einschließlich akuter myeloblastischer Leukämie, multiplem Myelom und hepatozellulärem Karzinom22,23.
Häufig verwendete biomolekulare In-vitro-Interaktionsnachweistechniken
Die Co-Immunpräzipitation (Co-IP) ist eine Technik, die sich auf die Spezifität von Antigen-Antikörpern stützt, um biologisch relevante Wechselwirkungen zu identifizieren24. Der Hauptnachteil dieser Methode ist ihre Unfähigkeit, Wechselwirkungen mit geringer Affinität und kinetische Werte zu erkennen24. Biophysikalische Methoden wie die isotherme Titrationkalorimetrie (ITC), die Oberflächenplasmonenresonanz (SPR), die Biolayer-Interferometrie (BLI) und die FEB-Technologie werden bevorzugt, um die kinetischen Werte zu bestimmen.
ITC ist eine biophysikalische Nachweismethode, die auf der Bestimmung der Bindungsenergie zusammen mit einer vollständigen thermodynamischen Analyse zur Charakterisierung biomolekularer Wechselwirkungenbasiert 25. Der Hauptvorteil von ITC besteht darin, dass keine Markierung oder Fixierung des Zielproteins erforderlich ist. Die Hauptschwierigkeiten, mit denen ITC konfrontiert ist, sind die hohe Konzentration des Zielproteins, die für ein Experiment erforderlich ist, und die Schwierigkeit, nicht-kovalente Komplexe aufgrund kleiner Bindungsenthalpienzu analysieren 26. Sowohl SPR als auch BLI sind markierungsfreie biophysikalische Techniken, die auf der Immobilisierung des Zielmoleküls auf der Sensoroberfläche beruhen, gefolgt von nachfolgenden Injektionen des Analyten über das immobilisierte Ziel27,28. Bei SPR werden Veränderungen des Brechungsindex während biomolekularer Wechselwirkungen gemessen27; In BLI wird die Interferenz im reflektierten Licht in Echtzeit als Änderung der Wellenlänge als Funktion der Zeit28 aufgezeichnet. Sowohl SPR als auch BLI haben gemeinsame Vorteile durch hohe Spezifität, Sensitivität und Erkennungsfunktionen29. Bei beiden Methoden wird das Zielprotein auf Biosensoroberflächen immobilisiert, und daher kann es zu einem gewissen Verlust der nativen Konformation des Ziels kommen, was es schwierig macht, zwischen spezifischen und unspezifischen Wechselwirkungenzu unterscheiden 30. BLI verwendet teure faseroptische Einweg-Biosensoren, um das Ziel zu immobilisieren, und ist daher eine kostspielige Technik31. Im Vergleich zu diesen etablierten biomolekularen Detektionswerkzeugen bietet die FEB-Technologie eine zuverlässige und markierungsfreie Plattform, indem sie niedrige nanomolare Konzentrationen für den biomolekularen Nachweis in Echtzeit mit kinetischer Charakterisierung verwendet. Die FEB-Technologie überwindet auch die sprudelnden Herausforderungen in der ITC und ist im Vergleich zu SPR oder BLI kostengünstiger.
Die auf dem Feldeffekttransistor (FET) basierenden Biosensoren sind ein aufstrebendes Gebiet zur Erkennung biomolekularer Wechselwirkungen, indem sie vielfältige biomedizinische Anwendungen anbieten. Im FET-System werden Ziele für die Biosensorchips immobilisiert und Wechselwirkungen durch Änderungen der Leitfähigkeit32 erkannt. Das Alleinstellungsmerkmal, das bei der Entwicklung eines effizienten elektronischen Biosensors zu berücksichtigen ist, sind die physikalisch-chemischen Eigenschaften wie die halbleitende Natur und die chemische Stabilität des Beschichtungsmaterials, das zur Herstellung der Sensoroberflächeverwendet wird 33. Herkömmliche Materialien wie Silizium, die für FET verwendet werden, haben die Empfindlichkeit von Sensoren eingeschränkt, da Oxidschichten zwischen dem Transistorkanal und einer spezifischen Umgebung für ein ordnungsgemäßes Funktionierenerforderlich sind 34. Darüber hinaus sind Siliziumtransistoren empfindlich gegenüber salzreichen Umgebungen, was es schwierig macht, biologische Wechselwirkungen in ihrer natürlichen Umgebung zu messen. Der graphenbasierte Biosensor wird als Alternative vorgestellt, da er eine hervorragende chemische Stabilität und ein elektrisches Feld bietet. Da Graphen eine einzelne Atomschicht aus Kohlenstoff ist, ist es sowohl als Halbleiter extrem empfindlich als auch chemisch mit biologischen Lösungen kompatibel; Beide Eigenschaften sind wünschenswert, um kompatible elektronische Biosensoren35 zu erzeugen. Das bemerkenswerte ultrahohe Belastungspotenzial von Biomolekülen, das graphenbeschichtete Biosensoren bieten, führte zur Entwicklung von graphenbasierten Biosensoren FEB-Technologie.
Prinzip der FEB-Technologie: FEB ist eine markierungsfreie biomolekulare Detektionstechnik, die den elektrischen Strom durch den Graphen-Biosensor misst, an den die Bindungsziele immobilisiert werden. Wechselwirkungen zwischen dem immobilisierten Protein und dem Analyten führen zu Stromänderungen, die in Echtzeit überwacht werden, was genaue kinetische Messungenermöglicht 36.
Instrumentierung: Das FEB-System besteht aus einem Graphen-Feldeffekttransistor (gFET)-Sensorchip und einem elektronischen Lesegerät, das während des gesamten Experiments eine konstante Spannung anlegt (Abbildung 1). Der Analyt wird in Lösung auf das Zielprotein aufgetragen, das auf der Biosensoroberfläche immobilisiert ist. Wenn eine Wechselwirkung auftritt, wird eine Veränderung des Stroms gemessen und in Echtzeit aufgezeichnet. Wenn die Analytkonzentration zunimmt, nimmt auch der Anteil des gebundenen Analyten zu, was zu höheren Veränderungen in der Strömung führt. Mit der mit dem Gerät gelieferten automatisierten Analysesoftware (Table of Materials) wird I-Response in Form von Biosensing-Einheiten (BU)37 gemessen und aufgezeichnet. I-Response ist definiert als die Veränderung des Stroms (I) durch den Biosensorchip, die in Echtzeit bei der Interaktion des immobilisierten Ziels mit dem Analyten gemessen wird. Die automatisierte Analysesoftware FEB kann sowohl die I-Response als auch die C-Response auf dynamische Interaktionsereignisse analysieren, wobei die C-Response die Änderungen der Kapazität (C) aufzeichnet. Die Variationen sowohl der I-Response als auch der C-Response entsprechen direkt dem Anteil des gebundenen Analyten und können weiter analysiert werden, um KD-Werte zu erzeugen. Die Standardeinstellung der automatisierten Analysesoftware ist I-Response.
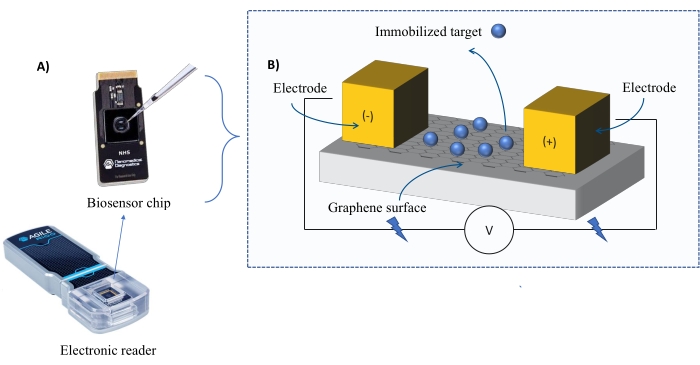
Abbildung 1: Überblick über den Versuchsaufbau . (A) Chip auf Graphenbasis und ein elektronisches Lesegerät. (B) Eine Übersicht über die Chipkomponenten. Der Chip ist an zwei Elektroden befestigt, die das System mit Strom versorgen. Die Oberfläche des Chips ist mit Graphen bedeckt, das bei Aktivierung das Ziel binden kann. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
Methodologie:
Zunächst wird der aktivierte Biosensorchip in das FEB-Gerät eingesetzt (Abbildung 1), gefolgt von der Ausführung der folgenden Schritte: (1) Kalibrierung: Das Experiment beginnt mit der Systemkalibrierung unter Verwendung von 1x phosphatgepufferter Kochsalzlösung (PBS; pH = 7,4), um die Ausgangsgleichgewichtsantwort zu erzeugen. (2) Assoziation: Der Analyt wird in den Chip eingeführt und die I-Antwort wird überwacht, bis die Bindungssättigung erreicht ist. (3) Dissoziation: Der Analyt wird mit 1x PBS dissoziiert. (4) Regeneration: Reste des Analyten werden mit 1x PBS entfernt. (5) Waschen: Insgesamt werden fünf Waschungen mit 1x PBS durchgeführt, um die gebundenen und ungebundenen Analyten gründlich vom Chip zu entfernen.
Analyse:
Die Datenanalyse erfolgt mit der vollautomatischen Software, die mit dem Gerät geliefert wird. Die automatisierte Analysesoftware generiert ein Hill-Fit-Diagramm mit einem KD-Wert . Das Hill-Fit-Diagramm beschreibt die Assoziation eines Analyten mit dem Zielprotein als Funktion der Analytkonzentrationen. Die Konzentration, bei der ein halbmaximales Ansprechen erreicht wird, ist proportional zum KD-Wert . Ein niedriger KD-Wert steht für eine hohe Bindungsaffinität und umgekehrt.
Um die aus dem FEB-Experiment gewonnenen Daten zu validieren, werden I-Responses von jedem Auslesepunkt für jede Analytkonzentration mit der Datenüberprüfungs-/Exportsoftware extrahiert und können in andere statistische Analysesoftware (siehe Tabelle der Materialien) exportiert werden, wie unten erläutert.
