Ziel dieser Methode ist es zu untersuchen, wie der retinale Input das Wachstum und die Entwicklung des optischen Tektums, des visuellen Verarbeitungszentrums im Zebrafischgehirn, beeinflusst. Durch Entfernen eines Auges und anschließenden Vergleich der beiden Seiten des optischen Tektums können tektale Veränderungen innerhalb derselben Probe beobachtet und normalisiert werden, was einen Vergleich zwischen mehreren Proben ermöglicht. Moderne molekulare Ansätze in Kombination mit dieser Technik werden Einblicke in die Mechanismen liefern, die dem Wachstum und der Entwicklung des visuellen Systems sowie der axonalen Degeneration und Regeneration zugrunde liegen.
Sensorische Systeme – visuelle, auditive und somatosensorische – sammeln Informationen von externen Organen und leiten diese Informationen an das zentrale Nervensystem weiter, wodurch “Karten” der äußeren Welt im Mittelhirn erzeugtwerden 1,2. Das Sehen ist die dominierende sensorische Modalität für fast alle Wirbeltiere, einschließlich vieler Fische. Die Netzhaut, das Nervengewebe im Auge, sammelt Informationen mit einem neuronalen Schaltkreis, der hauptsächlich aus Photorezeptoren, bipolaren Zellen und retinalen Ganglienzellen (RGCs), den Projektionsneuronen der Netzhaut, besteht. RGCs haben lange Axone, die ihren Weg über die innere Oberfläche der Netzhaut zum Sehnervenkopf finden, wo sie faszikulieren und gemeinsam durch das Gehirn wandern und schließlich im visuellen Verarbeitungszentrum im dorsalen Mittelhirn enden. Diese Struktur wird bei Fischen und anderen Nicht-Säugetierwirbeltieren als optisches Tectum bezeichnet und ist homolog zum Colliculus superior bei Säugetieren3.
Das optische Tectum ist eine bilateral symmetrische Mehrschichtstruktur im dorsalen Mittelhirn. Bei Zebrafischen und den meisten anderen Fischen erhält jeder Lappen des optischen Tektums visuelleEingaben ausschließlich vom kontralateralen Auge, so dass der linke Sehnerv im rechten Tektallappen und der rechte Sehnerv im linken Tektallappen 4 endet (Abbildung 1). Wie sein Gegenstück bei Säugetieren, der Colliculus superior, integriert das optische Tectum visuelle Informationen mit anderen sensorischen Eingaben, einschließlich Vorsprechen und Somatosensation, und steuert Verschiebungen der visuellen Aufmerksamkeit und Augenbewegungen wie Sakkaden 1,5,6. Im Gegensatz zum Colliculus superior des Säugetiers erzeugt das optische Tectum jedoch kontinuierlich neue Neuronen und Glia aus einer spezialisierten Stammzellnische in der Nähe der medialen und kaudalen Ränder der tektalen Lappen, der sogenannten tektalen Proliferationszone7. Die Aufrechterhaltung proliferativer Vorläufer im optischen Tectum und anderen Regionen des zentralen Nervensystems trägt zum Teil zu der bemerkenswerten Regenerationsfähigkeit bei, die in Zebrafisch8 dokumentiert ist.
Frühere Arbeiten, die das Gehirn von blinden oder einäugigen Fischen untersuchten, ergaben, dass die Größe des optischen Tektums direkt proportional zur Menge der retinalen Innervation ist, die sie erhält 9,10,11. Bei erwachsenen Höhlenfischen, deren Augen in der frühen Embryogenese degenerieren, ist das optische Tectum merklich kleiner als bei eng verwandten, gesichteten Oberflächenfischen9. Die Degeneration der Höhlenfischaugen kann blockiert werden, indem die endogene Linse während der Embryogenese durch eine Linse von einem Oberflächenfisch ersetzt wird. Wenn diese einäugigen Höhlenfische bis ins Erwachsenenalter aufgezogen werden, enthält der innervierte Tektallappen etwa 10% mehr Zellen als der nicht innervierte Tektallappen9. In ähnlicher Weise war bei Larvenkillifischen, die mit chemischen Behandlungen inkubiert wurden, um Augen unterschiedlicher Größe innerhalb desselben Individuums zu erzeugen, die Seite des Tektums mit mehr Innervation größer und enthielt mehr Neuronen10. Beweise aus Experimenten zur Quetschung des Sehnervs an erwachsenen Goldfischen deuten darauf hin, dass die Innervation die Proliferation fördert, wobei die Proliferation der tektalen Zellen abnimmt, wenn die Innervation gestört ist11.
Mehrere neuere Berichte, die diese klassischen Studien bestätigen und erweitern, liefern Daten, die darauf hindeuten, dass die Proliferation als Reaktion auf Innervation zumindest teilweise durch den BDNF-TrkB-Signalweg12,13 moduliert wird. Es bleiben viele offene Fragen zum Wachstum und zur Entwicklung des optischen Tektums, einschließlich der Frage, wie ein sich entwickelndes sensorisches System mit Verletzungen und Axondegeneration fertig wird, welche zellulären und molekularen Signale es ermöglichen, dass der Netzhauteintrag das Wachstum des optischen Tektums reguliert, wann diese Mechanismen aktiv werden und ob die innervationsbedingte Proliferation und Differenzierung es der Netzhaut und ihrem Zielgewebe ermöglicht, Wachstumsraten zu koordinieren und eine genaue retinotopische Kartierung zu gewährleisten. Darüber hinaus gibt es viel größere Fragen zur aktivitätsabhängigen Entwicklung, die durch die Befragung des visuellen Systems des Zebrafisches mit chirurgischen Ansätzen wie dem unten beschriebenen beantwortet werden können.
Um die zellulären und molekularen Mechanismen zu untersuchen, durch die neuronale Aktivität, insbesondere durch visuellen Input, das Überleben und die Proliferation von Zellen verändert, vergleicht der beschriebene Ansatz direkt innervierte und denervierte tektale Lappen (Abbildung 1) innerhalb einzelner Zebrafischlarven. Diese Methode ermöglicht die Dokumentation der RGC-Axondegeneration im optischen Tectum und die Bestätigung, dass die Anzahl der mitotischen Zellen mit der Innervation korreliert.
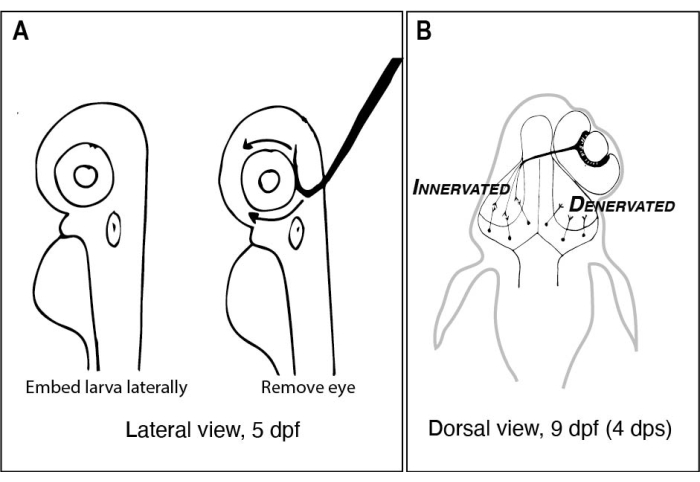
Abbildung 1: Skizzen von Zebrafischlarven vor und nach einseitiger Augenentfernung . (A) Zeichnung von 5 dpf-Larven unter einem Seziermikroskop. Jede Larve ist in Agarose mit niedrigem Schmelzpunkt eingebettet und seitlich ausgerichtet, bevor eine Wolframnadel mit einer scharfen, hakenförmigen Spitze verwendet wird, um das nach oben gerichtete Auge (in diesem Beispiel linkes Auge) herauszuholen. (B) Zeichnung der dorsalen Ansicht einer 9 dpf Larve, die aus der in A dargestellten Operation resultiert. Nur drei hochschematisierte RGC-Axone aus dem rechten Auge sind defaszikulierend und verbinden sich mit Neuronen im linken tektalen Lappen. Abkürzungen: dpf = Tage nach der Befruchtung; dps = Tage nach der Operation; RGC = retinale Ganglienzellen. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
