L’obiettivo di questo metodo è quello di indagare come l’input retinico influenza la crescita e lo sviluppo del tectum ottico, il centro di elaborazione visiva nel cervello del pesce zebra. Rimuovendo un occhio e quindi confrontando i due lati del tectum ottico, è possibile osservare e normalizzare i cambiamenti tectali all’interno dello stesso campione, consentendo il confronto tra più campioni. I moderni approcci molecolari combinati con questa tecnica forniranno informazioni sui meccanismi alla base della crescita e dello sviluppo del sistema visivo, nonché della degenerazione e rigenerazione assonale.
I sistemi sensoriali – visivi, uditivi e somatosensoriali – raccolgono informazioni dagli organi esterni e trasmettono tali informazioni al sistema nervoso centrale, generando “mappe” del mondo esterno attraverso il mesencefalo 1,2. La visione è la modalità sensoriale dominante per quasi tutti i vertebrati, compresi molti pesci. La retina, il tessuto neurale dell’occhio, raccoglie informazioni con un circuito neuronale costituito principalmente da fotorecettori, cellule bipolari e cellule gangliari retiniche (RGC), i neuroni di proiezione della retina. Gli RGC hanno lunghi assoni che trovano la loro strada attraverso la superficie interna della retina fino alla testa del nervo ottico, dove si fascicolano e viaggiano insieme attraverso il cervello, terminando infine nel centro di elaborazione visiva nel mesencefalo dorsale. Questa struttura è chiamata tectum ottico nei pesci e in altri vertebrati non mammiferi ed è omologa al collicolo superiore nei mammiferi3.
Il tectum ottico è una struttura multistrato bilateralmente simmetrica nel mesencefalo dorsale. Nel pesce zebra e nella maggior parte degli altri pesci, ogni lobo del tectum ottico riceve input visivi esclusivamente dall’occhio controlaterale, in modo tale che il nervo ottico sinistro termina nel lobo tectale destro e il nervo ottico destro termina nel lobo tectale sinistro4 (Figura 1). Come la sua controparte mammifera, il collicolo superiore, il tectum ottico integra le informazioni visive con altri input sensoriali, tra cui l’audizione e la somatosensazione, controllando i cambiamenti nell’attenzione visiva e nei movimenti oculari come le saccadi 1,5,6. Tuttavia, a differenza del collicolo superiore dei mammiferi, il tectum ottico genera continuamente nuovi neuroni e glia da una nicchia di cellule staminali specializzate vicino ai bordi mediali e caudali dei lobi tettoli chiamati zona di proliferazione tectale7. Il mantenimento dei progenitori proliferativi nel tectum ottico e in altre regioni del sistema nervoso centrale contribuisce, in parte, alla notevole capacità rigenerativa documentata nel pesce zebra8.
Lavori precedenti che esaminano il cervello di pesci ciechi o con un occhio solo hanno rivelato che la dimensione del tectum ottico è direttamente proporzionale alla quantità di innervazione retinica che riceve 9,10,11. Nei pesci delle caverne adulti, i cui occhi degenerano nell’embriogenesi precoce, il tectum ottico è notevolmente più piccolo di quello dei pesci di superficie strettamente imparentati e vedenti9. La degenerazione dell’occhio di pesce delle caverne può essere bloccata sostituendo la lente endogena con una lente da un pesce di superficie durante l’embriogenesi. Quando questi pesci delle caverne con un occhio solo vengono allevati fino all’età adulta, il lobo tectale innervato contiene circa il 10% in più di cellule rispetto al lobo tectale non innervato9. Allo stesso modo, nei killifish larvali che sono stati incubati con trattamenti chimici per generare occhi di dimensioni diverse all’interno dello stesso individuo, il lato del tectum con più innervazione era più grande e conteneva più neuroni10. Le prove degli esperimenti di schiacciamento del nervo ottico nel pesce rosso adulto indicano che l’innervazione promuove la proliferazione, con la proliferazione delle cellule tettoniche che diminuisce quando l’innervazione è stata interrotta11.
Confermando ed estendendo questi studi classici, diversi rapporti recenti forniscono dati che suggeriscono che la proliferazione in risposta all’innervazione è modulata, almeno in parte, dalla via BDNF-TrkB12,13. Rimangono molte domande aperte sulla crescita e lo sviluppo del tectum ottico, incluso il modo in cui un sistema sensoriale in via di sviluppo affronta la degenerazione di lesioni e assoni, quali segnali cellulari e molecolari consentono l’input retinico per regolare la crescita del tectum ottico, quando questi meccanismi diventano attivi e se la proliferazione e la differenziazione legate all’innervazione consentono alla retina e al suo tessuto bersaglio di coordinare i tassi di crescita e garantire un’accurata mappatura retinotopica. Inoltre, ci sono domande molto più grandi sullo sviluppo dipendente dall’attività che possono essere affrontate interrogando il sistema visivo del pesce zebra con approcci chirurgici come quello descritto di seguito.
Per studiare i meccanismi cellulari e molecolari con cui l’attività neurale, in particolare dall’input visivo, altera la sopravvivenza e la proliferazione cellulare, l’approccio descritto confronta direttamente i lobi tettoli innervati e denervati (Figura 1) all’interno delle singole larve di zebrafish. Questo metodo consente la documentazione della degenerazione dell’assone RGC nel tectum ottico e la conferma che il numero di cellule mitotiche è correlato all’innervazione.
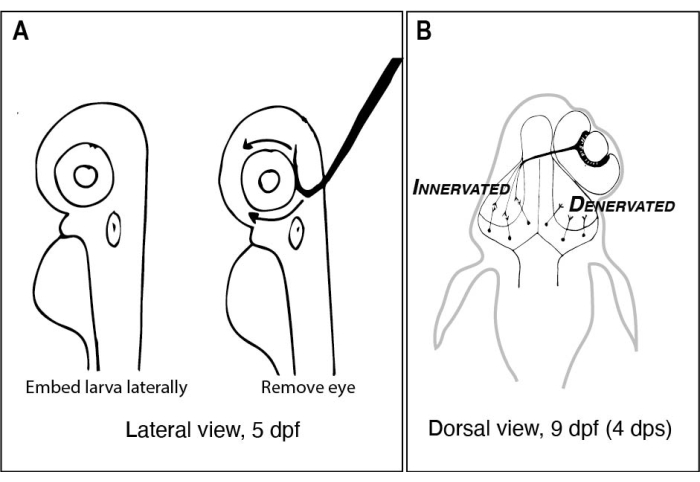
Figura 1: Schizzi di larve di zebrafish prima e dopo la rimozione unilaterale dell’occhio. (A) Disegno di 5 larve dpf viste al microscopio di dissezione. Ogni larva è incorporata in agarosio a basso punto di fusione e orientata lateralmente prima che un ago di tungsteno con una punta affilata e uncinata venga utilizzato per estrarre l’occhio rivolto verso l’alto (occhio sinistro in questo esempio). (B) Disegno della vista dorsale di una larva di 9 dpf risultante dall’intervento chirurgico raffigurato in A. Solo tre assoni RGC altamente schematizzati dall’occhio destro sono mostrati defascicolare e connettersi con i neuroni nel lobo tectale sinistro. Abbreviazioni: dpf = giorni dopo la fecondazione; dps = giorni dopo l’intervento; RGC = cellule gangliari retiniche. Fare clic qui per visualizzare una versione più grande di questa figura.
