Целью этого метода является исследование того, как вход сетчатки влияет на рост и развитие оптического тектума, центра визуальной обработки в мозге рыбок данио. Удалив один глаз, а затем сравнив две стороны оптического тектума, можно наблюдать и нормализовать тектальные изменения в одном образце, что позволяет сравнивать несколько образцов. Современные молекулярные подходы в сочетании с этой техникой дадут представление о механизмах, лежащих в основе роста и развития зрительной системы, а также аксональной дегенерации и регенерации.
Сенсорные системы – зрительная, слуховая и соматосенсорная – собирают информацию от внешних органов и передают эту информацию в центральную нервную систему, генерируя «карты» внешнего мира по среднему мозгу 1,2. Зрение является доминирующей сенсорной модальностью почти для всех позвоночных, включая многих рыб. Сетчатка, нервная ткань в глазу, собирает информацию с помощью нейронной цепи, состоящей в основном из фоторецепторов, биполярных клеток и ганглиозных клеток сетчатки (RGC), проекционных нейронов сетчатки. RGC имеют длинные аксоны, которые находят свой путь через внутреннюю поверхность сетчатки к головке зрительного нерва, где они фасцикулируют и путешествуют вместе через мозг, в конечном итоге заканчиваясь в центре визуальной обработки в спинном среднем мозге. Эта структура называется оптическим тектумом у рыб и других позвоночных, не являющихся млекопитающими, и гомологична верхнему колликулусу у млекопитающих3.
Оптический тектум представляет собой двусторонне симметричную многослойную структуру в спинном среднем мозге. У рыбок данио и большинства других рыб каждая доля зрительного тектума получает визуальный ввод исключительно от контралатерального глаза, так что левый зрительный нерв заканчивается в правой тектальной доле, а правый зрительный нерв заканчивается в левой тектальной доле4 (рисунок 1). Как и его аналог млекопитающих, превосходный колликулус, оптический тектум интегрирует визуальную информацию с другими сенсорными входами, включая прослушивание и соматозондирование, контролируя сдвиги в зрительном внимании и движениях глаз, таких как саккады 1,5,6. Однако, в отличие от верхнего колликулуса млекопитающих, оптический тектум непрерывно генерирует новые нейроны и глию из специализированной ниши стволовых клеток вблизи медиального и каудального краев тектальных долей, называемой зоной текальной пролиферации7. Поддержание пролиферативных предшественников в тектуме зрительного нерва и других областях центральной нервной системы частично способствует замечательной регенеративной способности, задокументированной у рыбок данио8.
Предыдущая работа, изучающая мозг слепых или одноглазых рыб, показала, что размер зрительного тектума прямо пропорционален количеству иннервации сетчатки, которую он получает 9,10,11. У взрослых пещерных рыб, глаза которых вырождаются в раннем эмбриогенезе, оптический тектум заметно меньше, чем у близкородственных, видимых поверхностных рыб9. Дегенерация глаза пещерной рыбы может быть заблокирована путем замены эндогенной линзы линзой от поверхностной рыбы во время эмбриогенеза. Когда эти одноглазые пещерные рыбы выращиваются до зрелого возраста, иннервированная тектальная доля содержит примерно на 10% больше клеток, чем неиннервированная тектальная доля9. Аналогичным образом, у личинок киллифиш, которые инкубировались с помощью химической обработки для создания глаз разных размеров в пределах одного и того же человека, сторона тектума с большей иннервацией была больше и содержала больше нейронов10. Данные экспериментов по раздавливанию зрительного нерва у взрослых золотых рыбок указывают на то, что иннервация способствует пролиферации, при этом пролиферация тектальных клеток уменьшается, когда иннервация была нарушена11.
Подтверждая и расширяя эти классические исследования, несколько недавних докладов предоставляют данные, свидетельствующие о том, что пролиферация в ответ на иннервацию модулируется, по крайней мере частично, путем BDNF-TrkB12,13. Остается много открытых вопросов о росте и развитии оптического тектума, в том числе о том, как развивающаяся сенсорная система справляется с травмой и дегенерацией аксонов, какие клеточные и молекулярные сигналы позволяют входу сетчатки регулировать рост зрительного тектума, когда эти механизмы становятся активными, и позволяют ли иннервационная пролиферация и дифференцировка сетчатке и ее целевой ткани координировать темпы роста и обеспечивать точное ретинотопическое картирование. Кроме того, существуют гораздо более широкие вопросы о развитии, зависящем от активности, которые можно решить, опросив зрительную систему рыбок данио с помощью хирургических подходов, таких как описанный ниже.
Чтобы исследовать клеточные и молекулярные механизмы, с помощью которых нейронная активность, в частности от визуального ввода, изменяет выживание и пролиферацию клеток, описанный подход непосредственно сравнивает иннервированные и денервированные тектальные доли (рисунок 1) в отдельных личинках рыбок данио. Этот метод позволяет документировать дегенерацию аксона RGC в тектуме зрительного нерва и подтвердить, что количество митотических клеток коррелирует с иннервацией.
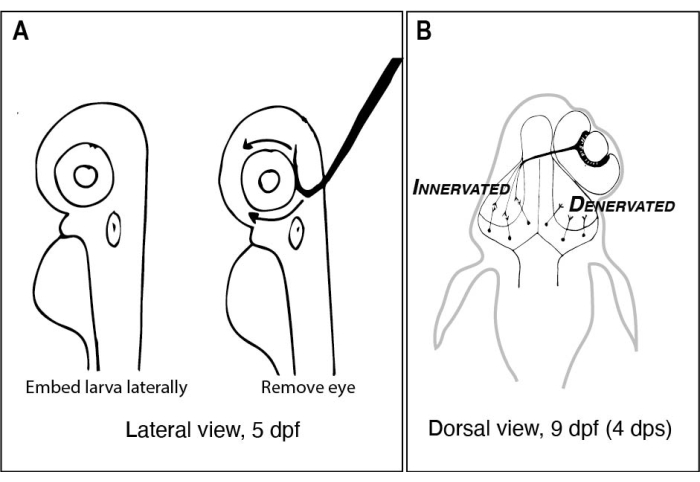
Рисунок 1: Эскизы личинок рыбок данио до и после одностороннего удаления глаз. (А) Рисование 5 личинок dpf при просмотре под рассекающим микроскопом. Каждая личинка встроена в агарозу с низкой температурой плавления и ориентирована сбоку, прежде чем вольфрамовая игла с острым крючковатым кончиком используется для вычерпывания глаза лицом вверх (левый глаз в этом примере). (B) Рисунок дорсального вида личинки 9 dpf в результате операции, изображенной в A. Только три сильно схематизированных аксона RGC от правого глаза дефасцикулируют и соединяются с нейронами в левой тектальной доле. Сокращения: dpf = дни после оплодотворения; dps = дни после операции; RGC = ганглиозные клетки сетчатки. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
